Lehe siseehitus sõltub küllalt tugevasti konkreetsetest ökoloogilistest tingimustest, eeskätt vee ja valguse kättesaadavusest. Kui uurida lehtede ehitust ühe taimeliigi isenditel, kes on kasvanud erineva tugevusega valguses, selgub mitmeid olulisi anatoomilisi iseärasusi. Taolisi muutusi nimetatakse kohanemisteks, sest need on mittepärilikud, pöörduvad muutused. Keskendume järgnevalt aga kohastumuslike, evolutsiooniliste muutuste vaatlemisele.
Veenõudluse alusel liigitatakse soontaimed neljaks:
1) mesofüüdid, taimed, kes on kohastunud kasvama humiidsetel aladel, kus temperatuur on mõõdukas ja mullaniiskus piisav, aga mitte liiga suur. Enamik Eesti taimeliike on mesofüüdid;
2) kserofüüdid ehk kuivustaimed, kelle ehituslikud ja talitluslikud kohastumused võimaldavad pikka aega taluda õhu ja mulla kuivust. Neil on kujunenud mitmesuguseid kohastumusi veevarude säilitamiseks ja säästlikuks kasutamiseks;
3) hügrofüüdid ehk niiskustaimed, kohastunud kasvama aladel, kus vett on mullas piisavalt ja veekulu suure õhuniiskuse tõttu väike;
4) hüdrofüüdid ehk veetaimed, kohastunud eluks vesikeskkonnas.
Lehelaba on kaetud elusatest rakkudest koosneva primaarse kattekoe -- epidermiga. Sõltuvalt asukohast eristatakse alumist (abaksiaalne ehk dorsaalne) ning ülemist (adaksiaalne ehk ventraalne) epidermi. Enamikul liikidest jääb epiderm ühekihiliseks, kuid esineb ka kahe-, kolme- ja enamakihilist epidermi. Epidermi lahutamatuks komponendiks on karvad (trihhoomid) ja õhulõhed, neil peatume allpool pikemalt.
Epidermirakud on enamasti lamedad, paksenenud väliskestaga. Kiiresti kasvavate taimeorganite epidermirakud on pikad ja kitsad, mõnedel muutlikes niiskustingimustes elavatel liikidel aga sopilised.
Enamasti jäävad epidermirakkude kestad tselluloosseteks ja rakud ise läbipaistvateks, kuid näiteks kõrrelistel (Poaceae), lõikheinalistel (Cyperaceae) ja osjadel (Equisetum) võivad rakukestad ränistuda. Välimisse rakukesta võib koguneda kaltsiumoksalaadi kristalle (draakonipuu -- Dracaena draco). Mõne sugukonna taimedel tekivad epidermirakkudes kaltsiumkarbonaadist kobarjad moodustised -- tsüstoliidid, mida peetakse kaitsekohastumuseks infektsiooni vastu.
Kloroplastid esinevad sõnajalgtaimede ja mõnede varjuliste kasvukohtade õistaimede epidermis ning enamusel taimedest õhulõhede sulgrakkudes. Viimastes võib leida ka värvituid ümaraid leukoplaste.
Epidermirakud eritavad väljapoole vaha ning kutiini - moodustub värvitu kile, kutiikula (cuticula). Kui vaha koguneb kutiikula pinnale (epikutikulaarne vaha), räägitakse vahakihist. Enamasti on see sinakashall kirme, nagu näiteks aednelgi (Dianthus caryophyllus) ja kapsa (Brassica oleracea) lehtedel. Mõnel taimel võib vahakiht olla õige paks, näiteks andi vahapalmil (Ceroxylon andicola) tüsedusega kuni 5 mm.
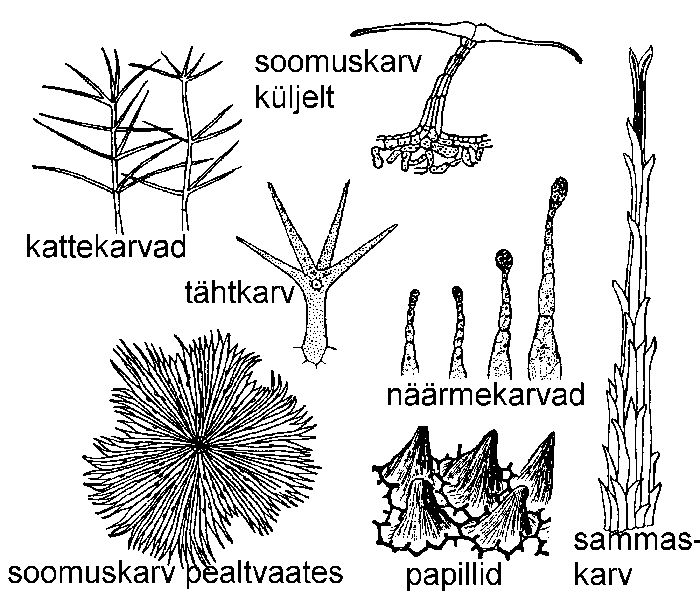
Epidermist võivad välja ulatuda väga mitmesuguse kuju ja suurusega, ühe- või mitmerakulised karvad ehk trihhoomid. Noored karvad on elusad, vanad, väljakujunenud karvad aga enamasti surnud.
Kui väljakasved jäävad madalateks väikesteks kühmudeks, nimetatakse neid papillideks ehk näsadeks. Papillid esinevad sagedasti õite kroonlehtedel, andes neile sametja välimuse.
Kattekarvad kaitsevad lehte intensiivse päikesekiirguse ja liigse transpiratsiooni eest. Täiskasvanult on need surnud, õhuga täitunud ja paistavad seetõttu valgete või hallidena. Lihtsamad kattekarvad on üherakulised liht- ja tähtkarvad, viimastel on karva ülaosa tähtjalt harunenud. Mitmerakulised karvad jagatakse üherealisteks, harunenuteks, soomus- ja sammaskarvadeks. Soomuskarv koosneb lühikesest jalast ja sellele kilpjalt kinnituvatest kitsastest rakkudest. Sammaskarv kujutab endast tihedasti üksteise vastu liibunud peente karvade kimpu. Olenevalt sellest, millise ilme annavad kattekarvad taimeosale, eristatakse siid-, ude-, taker-, vilt- ja karekarvu.
Haakekarvad on ühe- või mitmerakulised, tugevate rakukestadega. Neid leidub näiteks viljadel ja toele haakuvatel nõrkadel vartel.
Näärmekarvad koosnevad ühe- või mitmerakulisest jalast ja peast. Karvapea toodab mitmesuguseid eritusaineid.
Kõrvekarvad on enamasti üherakulised, kuid mitmerakulise alusega - subepidermaalsetest kudedest tekkinud emergentsi ehk väljakasvuga. Kõrvekarvade ehitust on kirjeldatud pikemalt töös nr. 65.
 Gaasivahetuseks on lehtedel erilised moodustised -- õhulõhed (stomata, ainsuses stoma).
Need tekivad meristemoidse õhulõhe emaraku
korduval jagunemisel. Harilikult paiknevad
õhulõhed lehel korrapäratult, harvem ridadena,
näiteks kitsaste lehtedega üheidulehelistel ja
okaspuudel. Õhulõhede kuju ja arv varieerub
liigiti olulisel määral.
Gaasivahetuseks on lehtedel erilised moodustised -- õhulõhed (stomata, ainsuses stoma).
Need tekivad meristemoidse õhulõhe emaraku
korduval jagunemisel. Harilikult paiknevad
õhulõhed lehel korrapäratult, harvem ridadena,
näiteks kitsaste lehtedega üheidulehelistel ja
okaspuudel. Õhulõhede kuju ja arv varieerub
liigiti olulisel määral.
Õhulõhe koosneb kahest sulgrakust, nende vahele jäävast õhupilust ning enamasti ka õhulõhe kaasrakkudest (neid on maksimaalselt seitse). Õhupilu välimist, laienenud osa nimetatakse esiõueks, sisemist tagaõueks, viimane laieneb õhukambriks -- õhulõhe all olevaks rakuvaheruumiks.
Õhulõhe on enamasti epidermiga ühes tasapinnas, aga võib ka eenduda (põhiliselt hüdrofüütidel) või paikneda epidermist sügavamal, süvendis (kserofüütidel). Kui ühte süvendisse avaneb mitu õhulõhet, nimetatakse seda koopaks ehk krüptiks.
Õhulõhed võivad asuda lehe mõlemal pinnal (amfistomaatiline leht) või ainult ühel. Kui õhulõhed esinevad ainult pealmisel pinnal, nimetatakse lehte epistomaatiliseks (näiteks veetaimede ujulehed), kui aga ainult alumisel pinnal, siis hüpostomaatiliseks (mitmed kserofüüdid).
Sõltuvalt kaasrakkude arvust eristatakse järgmisi õhulõhede tüüpe (joonis):
1) anomotsüütne -- kaasrakud puuduvad, näiteks jänesekapsas (Oxalis), võhumõõk (Iris);
2) anisotsüütne -- õhulõhed on ümbritsetud kolme kaasrakuga, millest üks on teistest märgatavalt väiksem, näiteks kukeharjal (Sedum);
3) paratsüütne -- kaasrakud paiknevad sulgrakkudega paralleelselt, näiteks kõrrelistel (Poaceae), tradeskantsial (Tradescantia);
 4) diatsüütne -- õhulõhel on kaks kaasrakku, mis asetsevad sulgrakkudega risti, näiteks nelgil (Dianthus);
4) diatsüütne -- õhulõhel on kaks kaasrakku, mis asetsevad sulgrakkudega risti, näiteks nelgil (Dianthus);
5) aktinotsüütne -- sulgrakud on ümbritsetud radiaalselt paiknevatest kaasrakkudest.
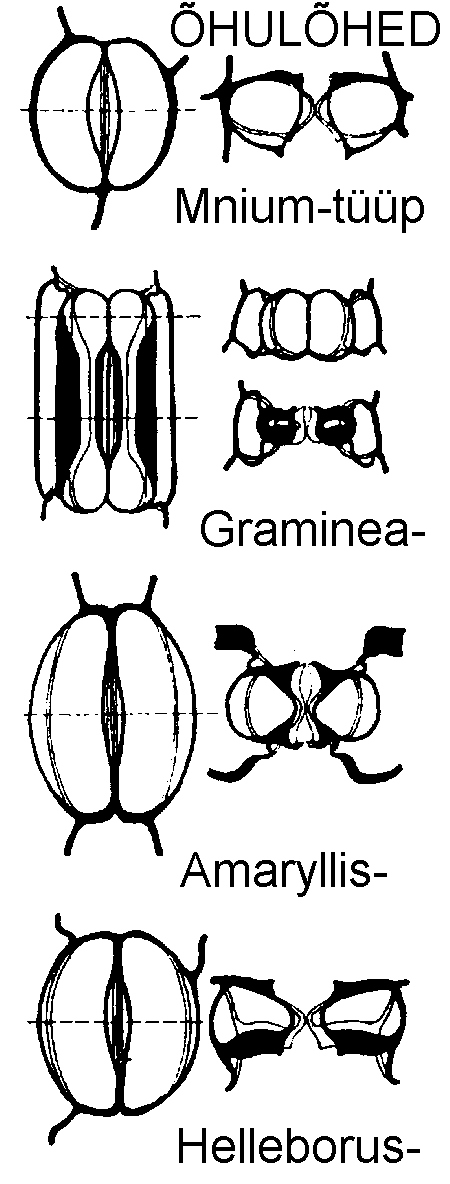
Teine õhulõhede klassifikatsioon põhineb sulgrakkude ehitusel. Selle järgi esineb nelja tüüpi õhulõhesid:
Mnium-tüüp (joonis). Õhupiluga piirnev rakukest on kõige õhem, teised kestad on enamasti paksenenud. Õhulõhe avanemise põhjustab sulgrakkude laienemine epidermi välispinnaga risti;
Amaryllis-tüüp. Sulgrakud on oakujulised. Õhupilu-poolne rakukest on ebaühtlaselt paksenenud, teistel külgedel on rakukest õhuke. Sulgrakud paisuvad epidermi pealmise pinnaga paralleelselt;
Helleborus-tüüp. Sarnaneb eelmisele, kuid sulgrakud liiguvad epidermi pealmise pinna suhtes samaaegselt igas suunas;
Graminea-tüüp. Sulgrakud on hantlikujulised, nende põisjad otsad on õhukesekestalised, keskosa aga jäik ja paksuseinaline. Rakkude otsimiste osade paisumisel eemalduvad rakkude keskmised osad üksteisest, turgori vähenemisel lähenevad taas. Seega liiguvad sulgrakud epidermi välimise pinnaga paralleelselt.
Kahe epidermi vahel olevat lehe põhikude nimetatakse mesofülliks. Kuna enamik lehe kloroplastidest paikneb mesofüllis, nimetatakse seda ka klorenhüümiks.
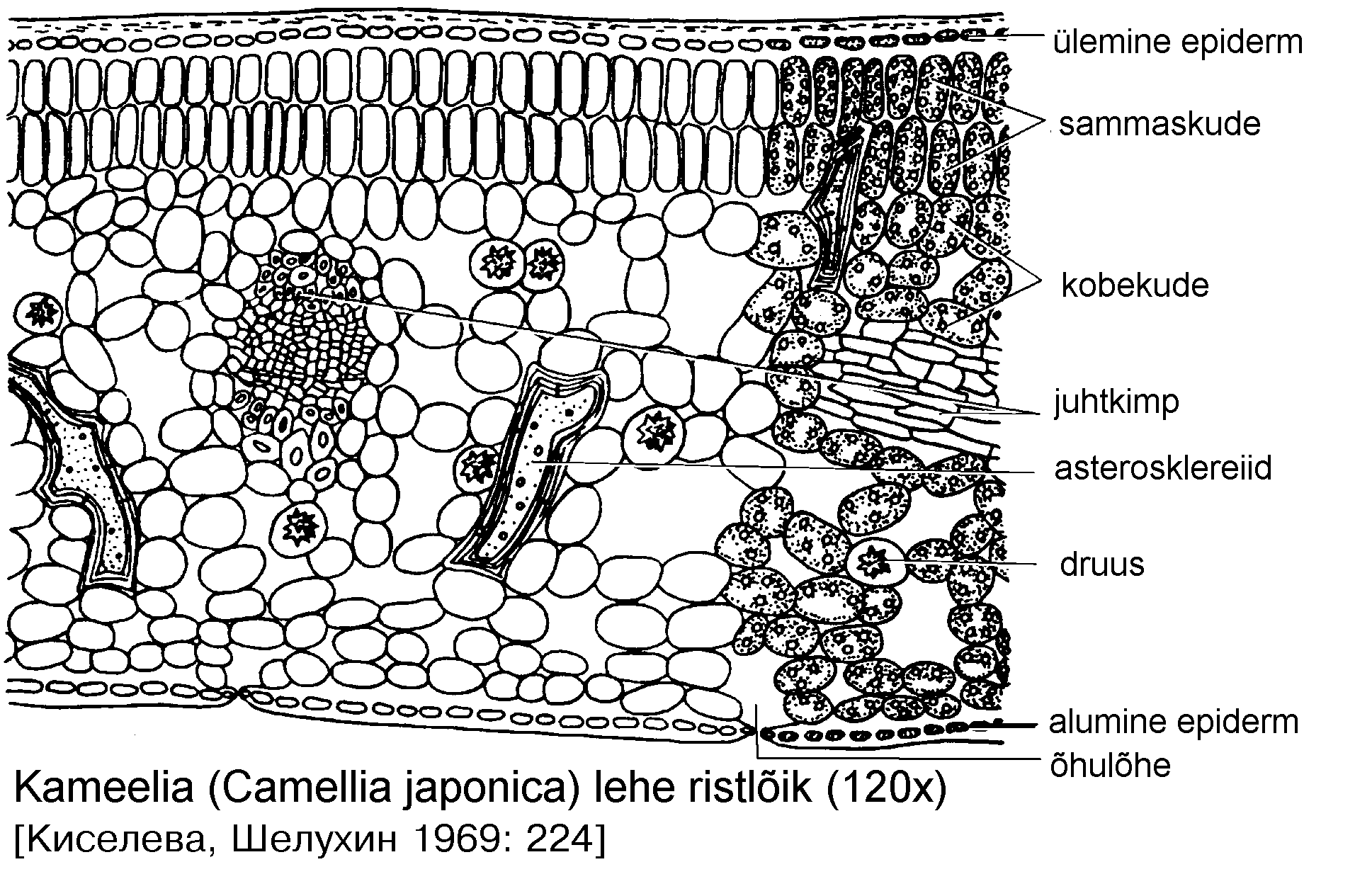
Mesofüll diferentseerub harilikult ühe või mitmekihiliseks sammaskoeks (palissaadkoeks) ning kobekoeks (tohlkoeks), kuid võib olla ka homogeenne.
Sammaskoe rakud asetsevad lehe välispinnaga risti. Enamasti koosneb sammaskude tihedalt asetunud piklikest rakkudest, mille vahele jäävad kitsad rakuvaheruumid. Vähediferentseerunud sammaskude ei tarvitse olla rangelt korrapärane. Parasvöötme taimedel areneb sammaskude reeglina ainult lehe ülemisel pinnal (dorsiventraalne leht). Kui sammaskude esineb lehe mõlemal pool, öeldakse, et leht on isolateraalne.
Sammas- ja kobekoe vahel on enamasti selge piir. Sisemised sammaskoe rakud toetuvad mõnekaupa kobekoe rakkudele, mida nimetatakse kogujarakkudeks.
Kobekoe rakkude suurus ja kuju on varieeruv, rakkude vahele jäävad mitmesuunalised rakuvaheruumid. Kobekude sisaldab tunduvalt vähem kloroplaste kui sammaskude, seepärast on enamik lehti pealt tumedamad kui alt.
 Üheidulehelistel taimedel ja okaspuudel on lehe tugikoeks ainult
sklerenhüüm, kaheidulehelistel sklerenhüüm ja kollenhümiseerunud
põhikude. Tugikudede asetus lehes võib olla väga mitmekesine. Vahel
leidub mesofüllis idioblastidena asterosklereiide (kameelia --
Camellia, teepõõsas -- Thea). Sageli esineb eeterlikke õlisid, vaike,
tanniine jms. eritavaid sekretoorseid rakke. Leidub ka kaltsiumoksalaadi kristalle.
Üheidulehelistel taimedel ja okaspuudel on lehe tugikoeks ainult
sklerenhüüm, kaheidulehelistel sklerenhüüm ja kollenhümiseerunud
põhikude. Tugikudede asetus lehes võib olla väga mitmekesine. Vahel
leidub mesofüllis idioblastidena asterosklereiide (kameelia --
Camellia, teepõõsas -- Thea). Sageli esineb eeterlikke õlisid, vaike,
tanniine jms. eritavaid sekretoorseid rakke. Leidub ka kaltsiumoksalaadi kristalle.
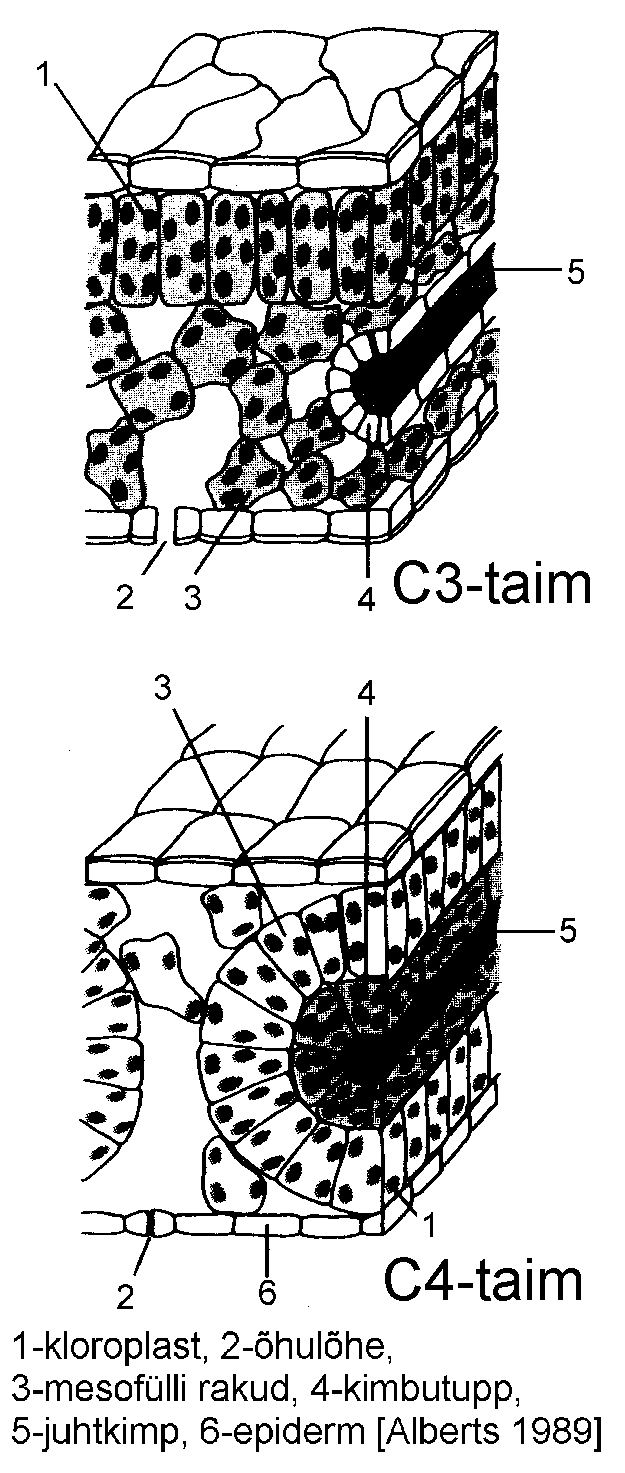
Mesofülli struktuur sõltub taime fotosünteesi tüübist. Enamik taimi on C3 tüüpi, nende lehe ehitus vastab ülalkirjeldatule. Soojematel aladel kasvab sageli C4 taimi (Eestis vaid üksikud liigid). Nende morfoloogiliseks iseärasuseks on mesofüllirakkude paiknemine tihedalt ümber juhtkimbu, selle tagajärjel moodustub nn. pärg-mesofüll (joonis).
Lehe ehitus sõltub olulisel määral ka kasvutingimustest. Isegi ühe ja sama isendi täiskasvanud lehed võivad olla erineva ehitusega. Puude võra välimised, nn. valguslehed on hästi arenenud sammaskoega, nende epidermirakud on paksema kestaga ning õhulõhesid on neil pinnaühiku kohta rohkem kui võra sees, halbades valgustingimustes kasvavatel varjulehtedel. Viimastel võib sammaskude olla ühekihiline või üldse puududa, tugikude on vähearenenud, epiderm õhem ning sisaldab kloroplaste.
Enamikul liikidel leidub mesofüllis erineva suurusega juhtkimpe, mida lehe puhul nimetatakse roodudeks. Need võivad olla kollateraalsed või bikollateraalsed, olenevalt sellest, millist tüüpi juhtkimbud esinevad varres. Lehes on bikollateraalsed vaid suuremad juhtkimbud.
Lehe juhtkimbud on enamasti kinnised (kambium esineb lühiajaliselt vaid arenevates lehtedes), harvem avatud (mõnedel igihaljastel taimedel, kuid kambium töötab neil nõrgalt ning sekundaarseid elemente juhtkimbus peaaegu ei ole).
Kõik juhtkimbud on ümbritsetud kimbutupega, mis koosneb põhikoelistest või sklerenhüümsetest rakkudest, harvem mõlemast korraga. Sarnaselt varte endodermile võivad mõne liigi kimbutupe rakud korgistuda.
Kimbutupe olemasolu tõttu ei puutu juhtkimbud rakuvaheruumidega vahetult kokku. Erandiks on hüdatoodid, mille ksüleem eritab epiteemi (modifitseerunud mesofülli) rakuvaheruumidesse vett. Vee gutatsioon (eritumine) lehest toimub vastava ava -- veelõhe (enamasti õhulõhe, mis on kaotanud sulgumisvõime) abil.
Kollateraalses lehe juhtkimbus asetseb ksüleem ülal ja floeem allpool. Juhtkimbu lõpp koosneb enamasti vaid ksüleemist, harvem ka floeemist. Peenemad juhtkimbud moodustavad lehes tavaliselt võrgustiku. "Võrgu silma", s.o. juhtkimpudega piiratud mesofülli osa nimetatakse areooliks. Areoolis võib leiduda juhtkimpude otsi.
Ka üheiduleheliste lineaalselt roodunud lehel moodustub mikroskoopilistest roodudest võrgustik. Niisuguste lehtede suuremaid paralleelselt paiknevaid juhtkimpe ühendavaid juhtkimbukesi nimetatakse kommissuurideks.
Leherootsu ehitus meenutab primaarse ehitusega vart. Juhtkimbud asetsevad selles kas ringina, laialipillatult või keskossa koondunult.
Heitlehiste puude igasügisesele lehelangemisele eelneb leherootsu ja oksa piiri kudedes rida anatoomilisi ja biokeemilisi muutusi, mille tulemusena lehe äralangemine ei kahjusta varre kudesid. Leherootsu ja varre piiril tekib eralduskiht, mille rakukestade lagunemine soodustab lehe eraldumist. Eralduskihist varre poole jääb korgistunud rakkudest moodustunud kaitsekiht, mis takistab transpiratsiooni ja parasiitide sissetungi. Varem või hiljem areneb kaitsekihist sügavamal periderm ning oks kattub ühtlase peridermikihiga. Samasugune eralduskiht tekib ka küpsete viljade või viljastamata õite varisemise eel. Senistel andmetel ei teki eralduskihti okaspuudel, näiteks kuusel (Picea sp.).
Kserofüütide (kuivustaimede) ehituslikud ja talitluslikud kohastumused võimaldavad neil aktiivsel elujärgul pikka aega taluda õhu ja mulla kuivust. Kuivustaimed kasvavad liivikutel, nõmmedel, kõrbetes jt. ariidsetel aladel. Kserofüüdid jaotatakse kolme rühma:
1) kseromorfsed taimed, kelle anatoomilised iseärasused, nagu paks kutiikula, tihe karvastik, lehtede kokkurullumine või reduktsioon jne. vähendavad transpiratsiooni. Sarnased kohastumused võivad kujuneda ka teiste ebasoodsate välistegurite (lämmastikuvaegus, ülemäärane kiirgus) toimel;
2) eukserofüüdid ehk päriskuivustaimed, nendel taimedel võib rakkude veesisaldus suurtes piirides kõikuda (tsütoplasma talub 50-80 % veekaotust), vee kerge kättesaadavuse korral tarbitakse seda ohtrasti;
3) turdtaimed ehk sukulendid, näiteks liigid sugukondadest kaktuselised (Cactaceae), piimalillelised (Euphorbiaceae), agaavilised (Agavaceae) jt. Nende lihakad, paksu kutiikula või vahakihiga kaetud varred on kohastunud vee säilitamiseks. Mesofüll on enamasti homogeenne, varre keskosa täidavad õhukesekestalised rakud, mis moodustavad veesäilituskoe.
Kserofüütide lehed on sageli pealt kurrulised, kaetud epidermi ja paksu kutiikulaga, viimast võib katta vahakiht. Mesofüütidega võrreldes on kserofüütidel reeglina rohkem õhulõhesid, kuid need on väiksemad. Karvadega kaitstud õhulõhed asuvad süvendites või lehe kurdude sisekülgedel. Veepuudusel rullub leht ülespoole kokku. Niimoodi on võimalik vähendada transpiratsiooni õhulõhesid päriselt sulgemata. Leht võib rulluda ka allapoole, näiteks sugukonnas kanarbikulised (Ericaceae), mürdilised (Myrtaceae). Õhulõhed asetsevad sel juhul lehe alumisel pinnal, karvade vahel.
Karvade kaitsefunktsiooni näiteks võib tuua vesi-kirburohu (Polygonum amphibium) vees (f. amphibium) ja maismaal (f. terrestre) kasvavad vormid. Esimese lehed on karvadeta, maismaavormil aga tihedalt karvased.
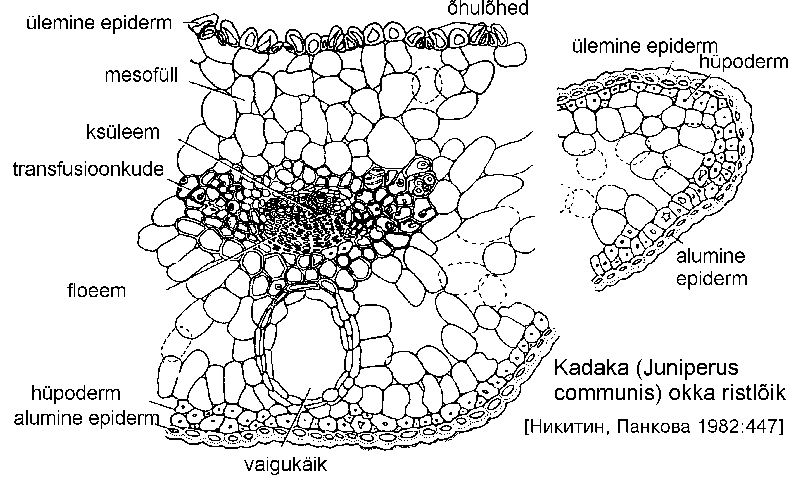
Okkad on tugevasti muundunud, kseromorfse ehitusega lehed. Läbilõikes on need kaarjad, rombjad või lamedad. Epidermirakkude paksenenud kestad on väljaspoolt kutiniseerunud.
Epidermi all asub sklerenhüümist või põhikoelistest rakkudest koosnev hüpoderm, mis vähendab kutikulaarset transpiratsiooni. Õhulõhed paiknevad kaasrakkudest moodustunud süvendites. Mesofüll on harilikult sammas- ja kobekoeks diferentseerumata. Selles esinevad vaigukäigud, mille arv ja pikkus erinevad perekonniti.
Okka keskel asuv steel on ümbritsetud paksukestalise endodermiga. Selle rakkudesse koguneb mõnikord tärklist ning kestades esineb Caspary jooni. Juhtkimpude naabruses paiknevad sklerenhüümirakud. Ülejäänud osa okka steelist on täidetud ülekande- ehk transfusioonkoega, mis koosneb koobaspooridega varustatud surnud ning üksikutest elusatest rakkudest. Surnud rakkude ülesanne on vee ja mineraalainete transport ksüleemist mesofülli, elusad rakud aitavad juhtida orgaanilisi aineid mesofüllist floeemi. Okka juhtkimbu sekundaarne kasv toimub väheaktiivse kambiumi talitlusel.
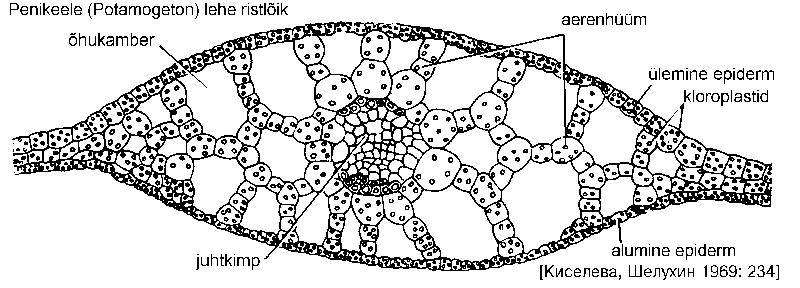
Veesiseste lehtede mesofüll on diferentseerumata. Hästi on aga arenenud rakuvaheruumistik ning õhukambrite võrk, neis säilitatakse elutegevuseks vajalikke gaase. Tänu õhukambrite olemasolule püsivad taimed vees vertikaalasendis. Gaasivahetus toimub kogu lehepinna (ja ka varre) kaudu, õhulõhed puuduvad või asetsevad ujulehtede pealmisel pinnal.
Veesisesed lehed on enamasti õhukesed ja koosnevad vaid mõnest rakukihist. Nende epiderm sisaldab kloroplaste. Lehtedel võib leiduda erilisi vahendeid vee imamiseks -- hüdropoode, näiteks kõõluslehel (Sagittaria sagittifolia) või vesikuusel (Myriophyllum). Hüdropoodide rakkude kestad lasevad vett kergesti läbi, kuna kutiikula neil puudub.
 Õistaimede kõige iseloomulikumaks organiks on õis
(flos). Kuna õie anatoomiale ja morfoloogiale pühendatud
uurimuste arv on aukartustäratav, puudub ühtne, enamiku
uurijate poolt aktsepteeritav õie käsitlus. Klassikalise
teooria kujunemist on oluliselt mõjutanud J. W. v. Goethe
filosoofilised ideed: õie homologiseerimine vegetatiivse
lühivõsuga, üksikuid õie osi aga lehtedega --
fülloomidega. Seetõttu esineb õie morfoloogia terminites
sageli sõna "leht", näiteks viljaleht või mikrosporofüll (kr.
phyllos -- leht).
Õistaimede kõige iseloomulikumaks organiks on õis
(flos). Kuna õie anatoomiale ja morfoloogiale pühendatud
uurimuste arv on aukartustäratav, puudub ühtne, enamiku
uurijate poolt aktsepteeritav õie käsitlus. Klassikalise
teooria kujunemist on oluliselt mõjutanud J. W. v. Goethe
filosoofilised ideed: õie homologiseerimine vegetatiivse
lühivõsuga, üksikuid õie osi aga lehtedega --
fülloomidega. Seetõttu esineb õie morfoloogia terminites
sageli sõna "leht", näiteks viljaleht või mikrosporofüll (kr.
phyllos -- leht).
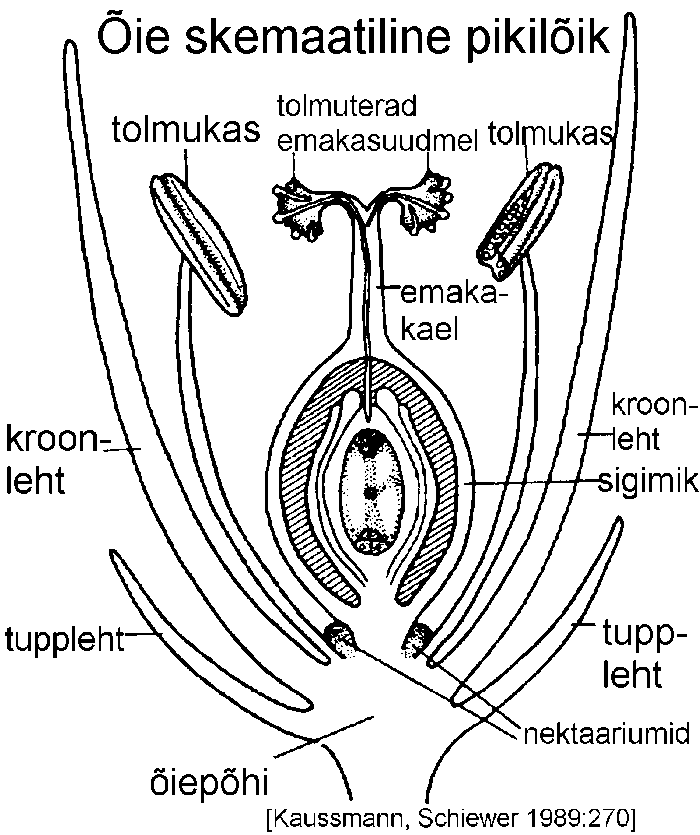
Õis koosneb fertiilsetest (ehk reproduktiivsetest) ning steriilsetest osadest, mis kõik kinnituvad õierao laienenud tipule -- õiepõhjale (receptaculum). Õieraag kinnitub varrele või õisiku puhul õisikuraole (joonis).
Õie steriilne osa, õiekate koosneb harilikult tupplehtedest, mis moodustavad tupe ning kroonlehtedest, mis moodustavad krooni. Paljude õistaimede puhul pole õiekatte selline jaotamine võimalik, sest õiekate koosneb ühesugustest lehtedest. Sellist lihtsat õiekatet nimetatakse perigooniks, vastandina tupeks ja krooniks jagunenud ehk kaheli õiekattele. Kui õiekate puudub, nimetatakse õit paljaks. Mitmesugustel õie osadel võib leiduda ekskretoorseid näärmeid -- nektaariume, mis eritavad suhkrurikast nektarit.
Õiekatte elemendid koosnevad põhikoest, juhtkimpudest ja epidermist, neis võib leiduda ka idioblaste, samuti kristallilisi moodustisi (rafiide või druuse) ja tärkliseteri. Kuigi rohelistes tupplehtedes esinevad kloroplastid, diferentseerub nende mesofüll sammas- ja kobekoeks väga harva. Nii tupp- kui kroonlehtedel võib esineda õhulõhesid. Õie steriilsete osade anatoomiline ehitus sarnaneb üldjoontes lehtede ehitusega.
Õies asuvad tolmukad moodustavad tolmukkonna (androeceum). Tolmukas (staminum) ehk mikrosporofüll on õie osa, kus arenevad tolmuterad. Enamasti on juba palja silmaga võimalik eristada tolmukapead (anthera) ja -niiti (filamentum). Nende ühinemiskohta nimetatakse konnektiiviks (connectivum) ehk nidemeks (joonis).
Tolmukaniidi anatoomiline ehitus on lihtne -- juhtkimpu ümbritseb põhikude, mida omakorda katab kutiniseerunud epiderm. Nii tolmukaniidil kui -peal võib esineda karvu ja õhulõhesid.
Tolmukapea koosneb kahest tolmukotist (saccus antherae). Ühes tolmukotis paikneb tavaliselt kaks tolmupesa ehk mikrosporangiumi (loculus antherae), sel juhul on tolmukas neljapesaline.
Vaheseina kadumisel võib tolmukott muutuda ühepesaliseks, näiteks käpaliste (Orchidaceae) sugukonnas. Mõne sugukonna taimedel, näiteks rööpkanarbikulistel (Epacridaceae), on vahesein tolmukottide vahelt kadunud, mistõttu kogu tolmukapead täidab vaid üks tolmupesa. Vaheseinte esinemisel võib tolmupesade arv ühes tolmukotis ka suureneda, näiteks liblikõieliste (Fabaceae) sugukonnas on tolmupesasid neli, puuvõõrikul (Viscum album) võib neid olla ligikaudu 50.
Tolmukad avanevad reeglina spontaanselt, vajamata selleks välist mõjurit. Tolmupesade vahele, kohta, kus endoteetsium puudub, tekib pikilõhe, mida nimetatakse stoomiumiks.
Kui lõhe tekib tolmukapea emakapoolsel küljel, nimetatakse sellist tolmukapead introorseks, kui aga vastasküljel, siis ekstroorseks. Avanemine võib toimuda ka ristilõhe, klappide ning teistsuguste avade abil, kuid teistsugused avanemisviisid esinevad pikilõhest harvemini.

1) epiderm ehk eksoteetsium (exothecium) -- ühekihiline;
2) endoteetsium ehk fibrooskiht (endothecium) -- suurte kiuliste kestadega ja kandilise kujuga, enamasti surnud rakkudest kiht;
3) vahekiht -- mitmekihiline, koosneb väikestest rakkudest, valminud tolmukapeas võib olla kadunud;
4) tapetkiht (tapetum) -- ühekihiline, väikestest rakkudest, valminud tolmukapeas võib samuti olla kadunud.
Osal sugukondadest koosneb tapetkiht suurest amöboidsest, hulgatuumalisest rakust ehk periplasmoodiumist, mis on tekkinud paljude rakkude liitumisel pärast nende kestade kadumist. Erinevalt sellisest amöboidsest tapetkihist koosneb sekretoorne tapetkiht, mis esineb eelmisest sagedamini, tugevasti arenenud endoplasmaatilise retiikulumi ja diktüosoomidega rakkudest. Rakud lagunevad pärast tolmuterade teket, nende kestad lahustuvad ning tsütoplasma satub tolmukapesasse, kus seda kasutatakse tolmutera kestade ülesehitamisel. Mõnedes sugukondades tarvitatakse tolmuterade kasvuks ka vahekihti talletatud ühendeid. Sel juhul võib vanades tolmukapeades vahekiht olla hävinud.
Tolmupesades paikneb õietolm, mis koosneb tolmuteradest (pollen). Tolmutera on homoloogne sõnajalgtaimede isasgametofüüdiga. Tolmuterade suurus on keskmiselt 15-50 ĩm, kuju võib olla kerajas, ovaalne, piklik või kuupjas. Tolmuterade arv sõltub liigist ja tolmlemisviisist - eriti ohtralt tekib tolmuteri tuultolmlejatel taimedel.
Tolmuteradel on kaks kesta: sisemine intiin ja välimine eksiin. Hügroskoopne ja tugevasti valgust murdev intiin võib olla ühe- või kahekihiline. Eksiin on mitmekihiline, selles leidub poore ja vagusid, mida koos nimetatakse apertuuriks (joonis). Enamasti kasvab tolmutoru just apertuuri kaudu välja. Madalama arengutasemega taimedel moodustavad apertuuri vaod, kõrgemalt arenenutel - poorid. Poore on 1-40, enamasti siiski 1-3. Eksiin on sageli liigispetsiifiliselt struktureerunud. See sisaldab lisaks tselluloosile ja kutiinile kõdunemisele väga vastupidavat sporopolleniini. Intiin sisaldab tselluloosi ja pektiini, säilitusainetest tärklist, suhkruid, valke ja rasvu.
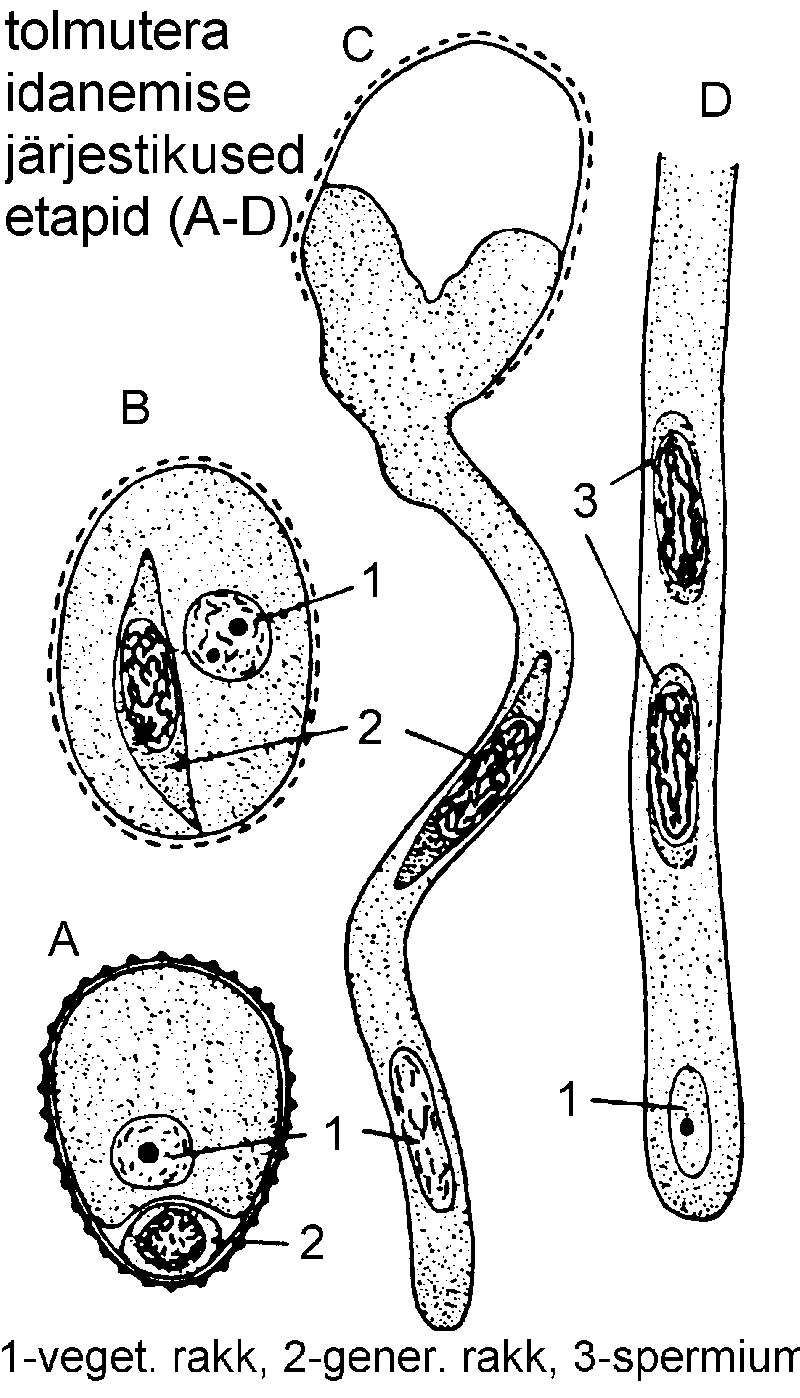
Tolmutera areng koosneb järgmistest etappidest.
1. Arhesporogenees. Noores arenevas tolmukapeas jaguneb üks subepidermaalne meristemaatiline rakk periklinaalselt. Sisemine rakk suureneb ning muutub arhespooriks, välimine jaguneb korduvalt, andes endoteetsiumi, vahe- ja tapetkihi.
2. Mikrosporogenees. Arhespoor jaguneb, moodustades suure hulga tolmuterade emarakke (mikrosporotsüüte). Iga tolmutera emarakk jaguneb meiootiliselt, andes neli haploidset mikrospoori, mis moodustavad ühe tetraadi.
Mikrospoorid võivad tekkida kas suktsessiivselt (igale tuuma jagunemisele järgneb kohe vahekestade moodustumine ja uute rakkude teke) või simulatiivselt (rakukestad moodustuvad korraga pärast tuumade jagunemise lõppemist). Esimene moodus on enam levinud üheidulehelistel, teine aga kaheidulehelistel taimedel.
Järgnevalt tetraad harilikult laguneb, kuid näiteks kanarbikuliste (Ericaceae) sugukonnas jäävad mikrospoorid kokku. Mikrospoorid kattuvad kestadega ning siitpeale nimetatakse neid juba tolmuteradeks.
3. Mikrogametogenees. Tolmutera tuum jaguneb mitootiliselt, moodustades vegetatiivse (sifonogeense) ja generatiivse (spermiogeense) raku. Esimene neist täidab enamiku tolmuterast. Kui tolmutera satub emakasuudmele või vastavasse toitekeskkonda, moodustab vegetatiivne rakk tolmutoru. Generatiivne rakk annab mitootiliselt jagunedes kaks spermiumi (isasgameeti). Spermiumid moodustuvad tavaliselt siis, kui tolmutera asub tolmukotis, näiteks sugukondades korvõielised (Asteraceae) ja kõrrelised (Poaceae). Sel juhul on valminud tolmutera kolmerakuline. Mõnes taimeperekonnas, nagu liilia (Lilium), tubakas (Nicotiana) või tradeskantsia (Tradescantia) lõppeb tolmutera areng generatiivse raku moodustumisega, sel juhul jäävad tolmuterad emakasuudmele sattumiseni kaherakulisteks ning spermiumid tekivad alles tolmutorus.
Kõrvalekaldeid normaalsest tolmutera arengust võivad põhjustada ebasoodsad ilmastikutingimused, mitmesugused keemilised ühendid ning geneetilised põhjused (hübriidsus, polüploidsus, apomiktilisus). Need viivad steriilse õietolmu tekkimisele.
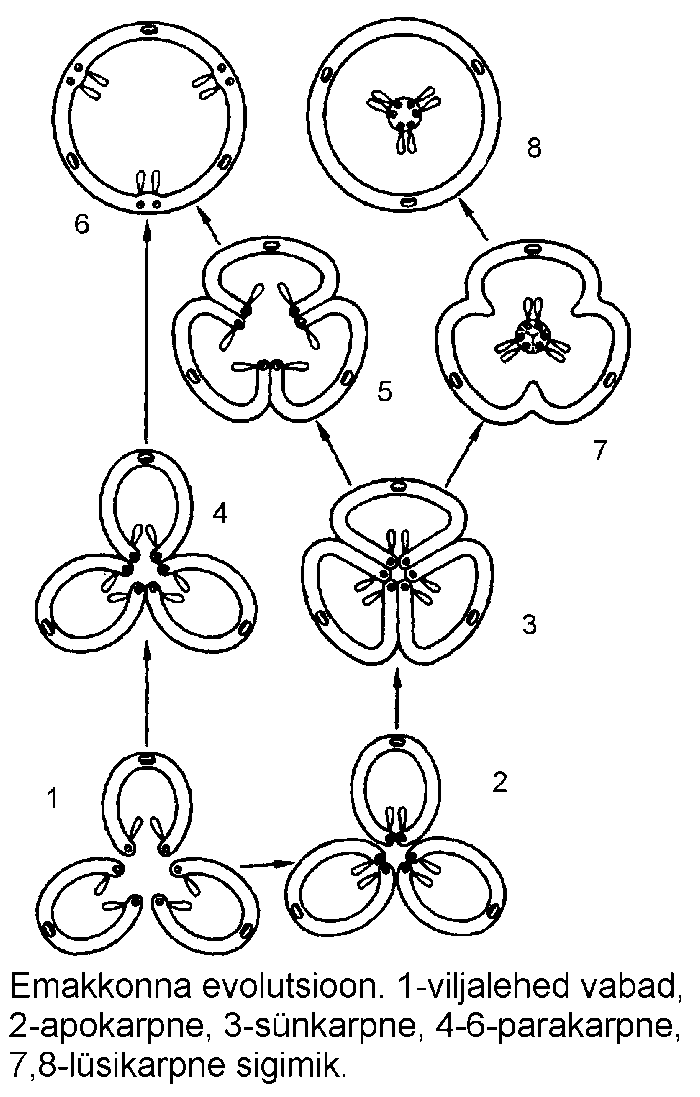 Ühe õie emakad moodustavad emakkonna
(gynoeceum). Emakas (pistillum) koosneb ühest
või mitmest viljalehest (carpellum). Kahe või
enama viljalehe esinemisel võivad need olla kas
vabad (lahkviljalehine ehk apokarpne sigimik) või
liitunud (liitviljalehine ehk sünkarpne sigimik)
(joonis). Ühest viljalehest koosnevat emakat käsitletakse apokarpsena.
Ühe õie emakad moodustavad emakkonna
(gynoeceum). Emakas (pistillum) koosneb ühest
või mitmest viljalehest (carpellum). Kahe või
enama viljalehe esinemisel võivad need olla kas
vabad (lahkviljalehine ehk apokarpne sigimik) või
liitunud (liitviljalehine ehk sünkarpne sigimik)
(joonis). Ühest viljalehest koosnevat emakat käsitletakse apokarpsena.
Emaka puhul saab harilikult eristada sigimikku (ovarium), emakakaela (stylus) ja -suuet (stigma). Kui emakakael on redutseerunud, nimetatakse vahetult sigimikule kinnituvat emakasuuet istuvaks.
Sigimik koosneb seinast ja sigimikupesast, mitme viljalehe olemasolul esinevad ka vaheseinad. Sigimikupesade arvu järgi jaotatakse sigimikud
1) ühepesalisteks, näiteks kirburohul (Polygonum sp.) või oblikal (Rumex sp.);
2) kahepesalisteks, näiteks harilikul vahtral (Acer platanoides);
3) kolmepesalisteks, näiteks liilialiste (Liliaceae) või amarülliliste (Amaryllidaceae) sugukonnas;
4) neljapesalisteks, näiteks kaheaastasel kuningakepil (Oenothera biennis);
5) viiepesalisteks, näiteks hibiskil (Hibiscus sp.);
6) paljupesalisteks, näi-teks vesikupul (Nuphar sp.);
7) ebatäielikult palju-pesalisteks, kui vaheseinad lõpevad enne sigimiku keskkohta, näiteks magunal (Papaver sp.).
Seda osa sigimikust, millele kinnituvad seemnealgmed, nimetatakse platsentaks (placenta). Igal viljalehel on kaks platsentat. Johtuvalt seemnealgmete kinnitumiskoha asendist ehk platsentatsioonist (placentatio), eristatakse apokarpses sigimikus
marginaalset platsentatsiooni, kui seemnealgmed kinnituvad viljalehe servadele, ning
laminaalset platsentatsiooni, kui seemnealgmed kinnituvad viljalehe keskele. Viimast peetakse primitiivsemaks.
Sünkarpses sigimikus võib platsentatsioon olla kas
 parietaalne, kui seemnealgmed kinnituvad
sigimiku seinale nendes kohtades, kus viljalehed on kokku kasvanud (parakarpses
sigimikus) või tsentraalne, kui seemnealgmed kinnituvad sigimiku keskosas
olevale sambale (lüsikarpne sigimik).
parietaalne, kui seemnealgmed kinnituvad
sigimiku seinale nendes kohtades, kus viljalehed on kokku kasvanud (parakarpses
sigimikus) või tsentraalne, kui seemnealgmed kinnituvad sigimiku keskosas
olevale sambale (lüsikarpne sigimik).
Platsentatsioonitüübi hõlpsamaks tuvastamiseks uuritakse juhtkimpude asetust viljalehes. Reeglina on ühel viljalehel kolm roodu -- keskmine ja kaks külgmist. Seemnealgmetesse suunduvad juhtkimbud algavad harilikult külgroodudest. Tsentraalse platsentatsiooni puhul paiknevad külgmised juhtkimbud sigimiku keskel, floeem seespool ja ksüleem väljaspool.
Seemnealgmeid on igas seemnepesas kas
üks, näiteks kirburohul (Polygonum sp.);
kaks, näiteks madaral (Galium sp.);
kolm, näiteks piimalillel (Euphorbia sp.) või
mitu, näiteks magunal (Papaver sp.).
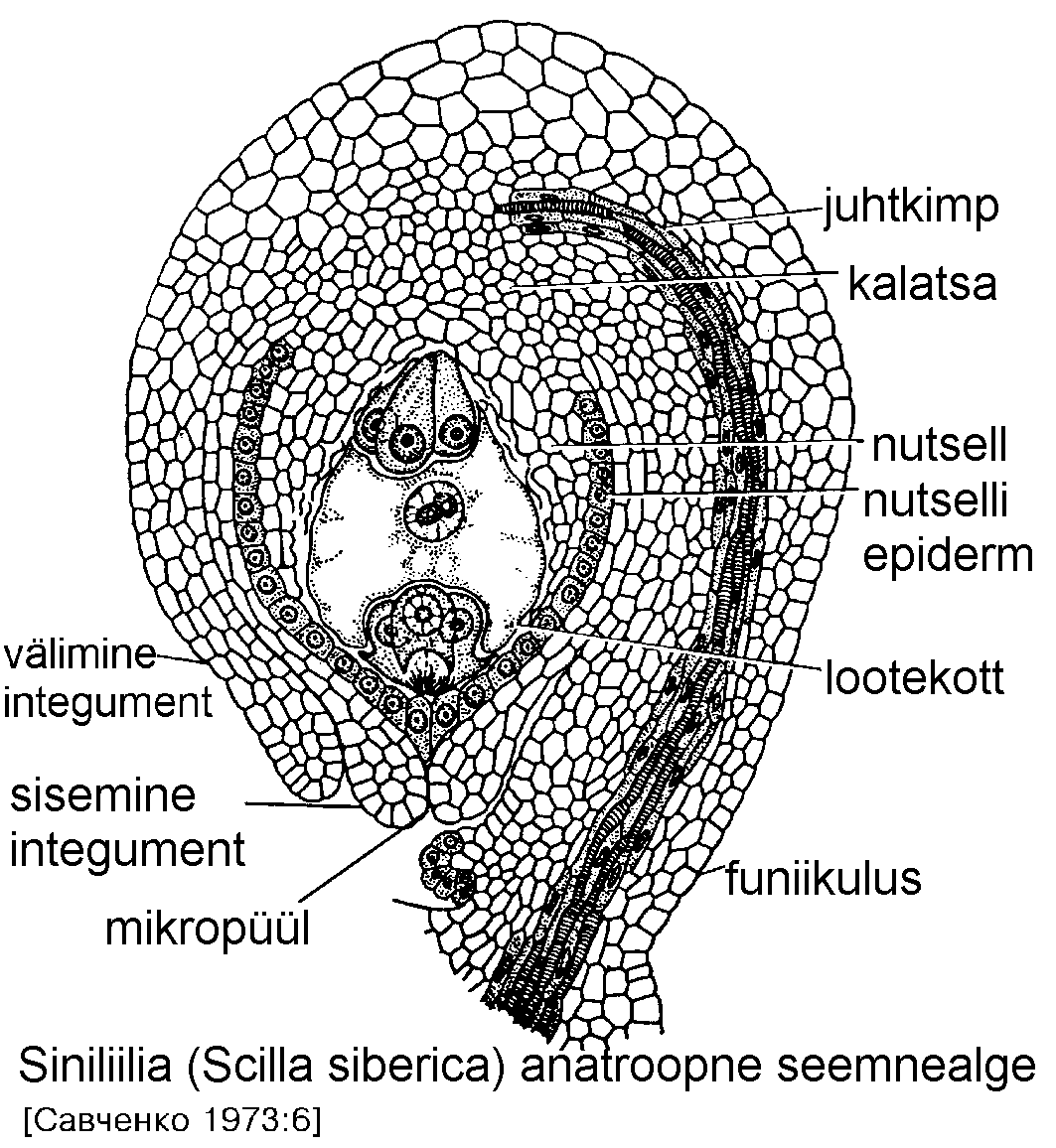
Seemnealgmed kinnituvad platsentale ning saavad toitained seemnealgme jala ehk funiikuluse (funiculus) kaudu (joonis). Kohta, kus lõpeb seemnealgme jalg ja selle juhtkimp ning algab seemnealge, nimetatakse kalatsaks.
Seemnealget katab üks või kaks integumenti ehk katet (välimine ja sisemine). Integumendid on epidermaalset päritolu, seetõttu võib neis eristada kolme kutiikula kihti -- välimine katab välimist integumenti, keskmine on integumentide vahel ning sisemine paikneb nutselli ja sisemise integumendi vahel. Seemnealgme tipul on integumentide vahel ava -- mikropüül. Viljastamisel tungib tolmutoru seemnealgmesse reeglina just siit.
 Seemnealgme sisemine
osa on nutsell ehk tuumik. Lootekoti arengus
võib nutsell täielikult
kaduda. Sel juhul muutub
integumendi sisekiht
rikkaliku endoplasmaatilise retiikulumiga
integumentaalseks tapetkihiks ehk endoteeliks.
Seemnealgme sisemine
osa on nutsell ehk tuumik. Lootekoti arengus
võib nutsell täielikult
kaduda. Sel juhul muutub
integumendi sisekiht
rikkaliku endoplasmaatilise retiikulumiga
integumentaalseks tapetkihiks ehk endoteeliks.
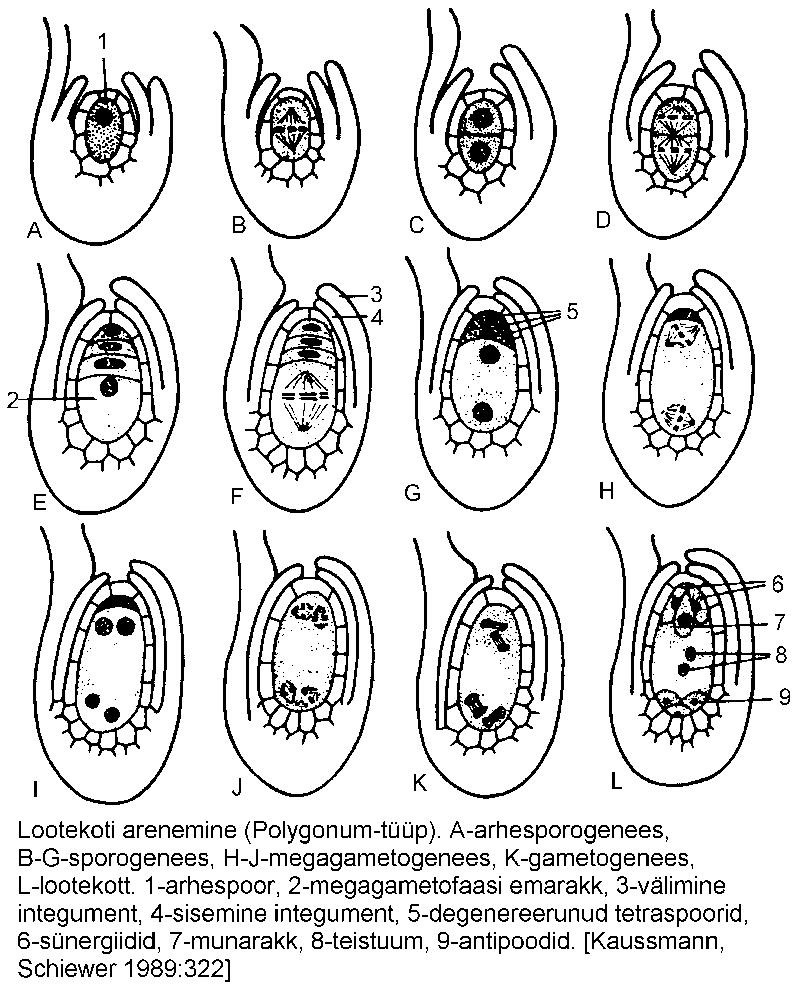
Mikropüüli ja platsenta vastastikuse asendi alusel eristatakse kaht seemnealgme põhitüüpi -- atroopne (ortotroopne) ehk püstine, kus mikropüül asub platsentast vastasküljel ning anatroopne ehk tagasikäändunud (joonis), kus mikropüül paikneb platsentaga ühel küljel. Teised tüübid on nende vahepealsed vormid (joonis). Munaraku tüüpiline arenguskeem ehk Polygonum-tüüpi lootekoti areng (joonis) koosneb järgmistest etappidest:
1) arhesporogenees -- seemnealgme nutselli mõned rakud suurenevad ning muutuvad arhespoorideks;
2) sporogenees -- arhespoorne diploidne rakk jaguneb meiootiliselt, andes neli ridastikku asetunud haploidset rakku ehk makrospoori. Kolm (tavaliselt ülemist) degenereeruvad, alumine (kalatsapoolne) suureneb ning jääb megagametofaasi emarakuks;
3) megagametogenees -- emaraku tuum jaguneb kolm korda, moodustades suure, kaheksatuumalise raku -- lootekoti;
4) gametogenees -- lootekott diferentseerub rakkudeks. Munarakku ümbritseb kaks väiksemat rakku - sünergiidi. Lootekoti kalatsapoolses osas on samuti kolm rakku - antipoodi, mis on seotud lootekoti toitumise ja kasvamisega. Kaks ülejäänud rakku, polaartuuma, liiguvad lootekoti keskossa ning ühinevad diploidseks teistuumaks. Lootekotis on seega kokku seitse rakku.
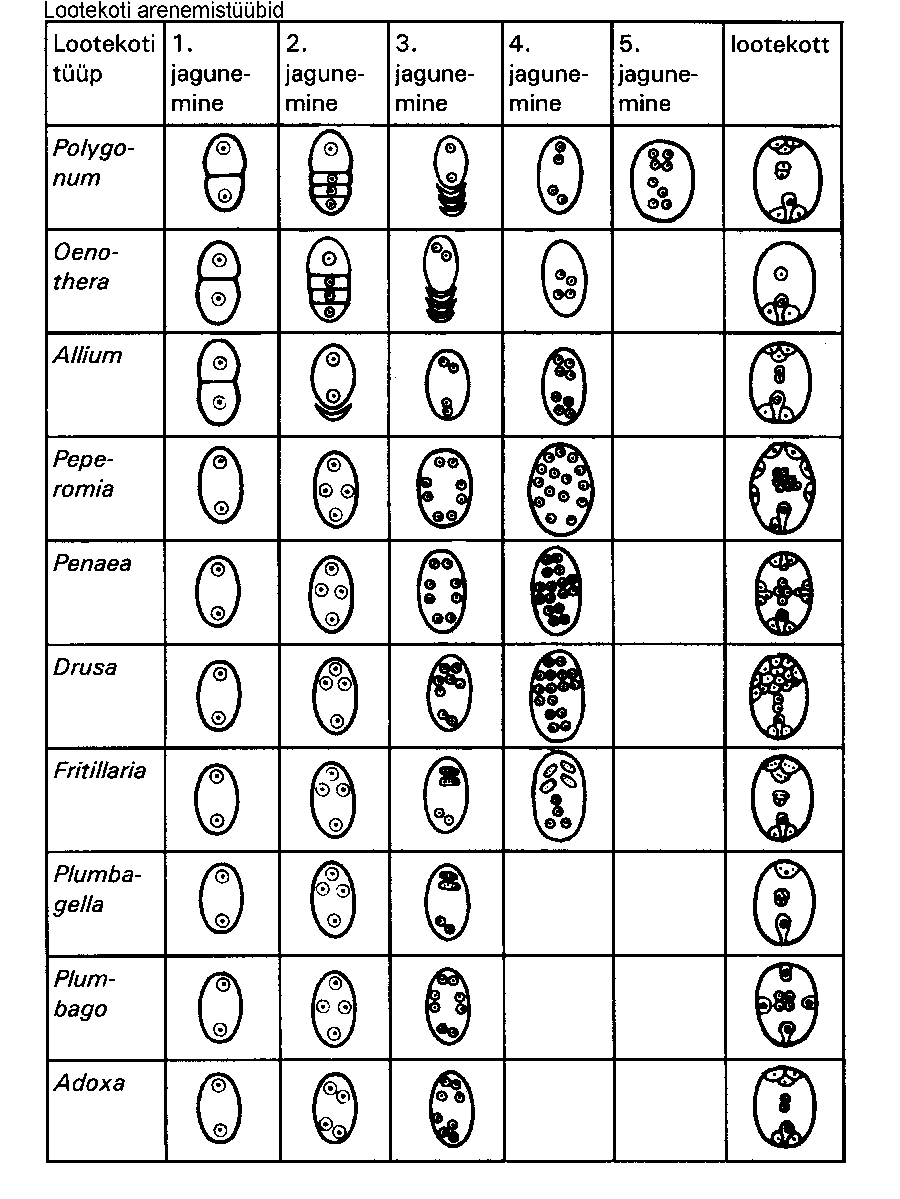
Polygonum-tüüp esineb suurel osal õistaimedest. Lootekoti teised arengutüübid on näidatud tabelis (joonis). Need grupeeritakse järgmiselt:
1) monospoorsed lootekotid (Polygonum ja Oenothera tüüp) -- lootekott moodustub makrospooride tetraadi ühest rakust.
2) bispoorsed lootekotid (Allium-tüüp) -- lootekott moodustub makrospooride diaadi ühest rakust.
3) tetraspoorsed lootekotid (kõik ülejäänud tüübid) -- lootekott moodustub neljatuumalisest makrospoorist, sest tuumi eraldavaid rakukesti ei teki, seega ei moodustu ka diaade ja tetraade.
Taimedel esineb sageli normaalsest munaraku arengust mitmesuguseid kõrvalekaldeid kas looduslike või kunstlike mõjutuste tagajärjel. Ka haploidsetel, hübriidsetel ja polüploidsetel taimedel on munaraku areng teistsugune. Sõltumata tekkepõhjustest viivad kõrvalekalded steriilse või vähearenenud lootekoti moodustumiseni.
Munaraku viljastamisele järgneb idu ja seemne arenemine, sigimikust moodustub aga vili (fructus). Õiekatted ning tolmukad kuivavad ja langevad ära, emakakael kas kuivab või muutub vilja levimise abivahendiks (haakekonksuks vms.).
Vili ei tarvitse alati koosneda ainult edasiarenenud sigimikust, mitmetes sugukondades esineb rüüsvilju (vananenud nimetus - ebavili), kus viljade arenemisest võtab lisaks viljalehtedele osa ka teisi kudesid, näiteks õunvilja või maasika lihakas osa on arenenud õiepõhjast. Levinud nähtuseks on viljakestade kokkukasvamine seemnekestadega, mis muudab viljad väliselt seemnetega sarnaseks. Tihti esineb ka partenokarpiat -- seemneteta vilja arenemist. Partenokarpia tekib osal liikidel spontaanselt, välismõjuta, kuid teistel on selleks vaja, et õied tolmleksid võõra õietolmuga (indutseeritud partenokarpia).
 Viljasein ehk perikarp võib olla arenenud ainult emaka
seinast või ka teistest kudedest. Kui perikarp koosneb
erineva anatoomiaga kihtidest, kasutatakse võrdlevas
morfoloogias nende kirjeldamisel mõisteid eksokarp,
mesokarp ja endokarp. Ontogeneetiline ja evolutsiooniline
tähendus neil mõistetel puudub.
Viljasein ehk perikarp võib olla arenenud ainult emaka
seinast või ka teistest kudedest. Kui perikarp koosneb
erineva anatoomiaga kihtidest, kasutatakse võrdlevas
morfoloogias nende kirjeldamisel mõisteid eksokarp,
mesokarp ja endokarp. Ontogeneetiline ja evolutsiooniline
tähendus neil mõistetel puudub.
Viljade morfoloogilistes klassifikatsioonides lähtutakse õie ja sigimiku ehitusest. Vili tekib ühest sigimikust, mis võib koosneda paljudest viljalehtedest. Koguvili moodustub apokarpsest sigimikust, kus viljalehed küpses viljas ei liitu (vaarikas -- Rubus idaeus). Vilikond areneb tervest õisikust, s.o. paljude õite viljumisel. Kui viljad arengu käigus liituvad, tekib liitvilikond, näiteks mooruspuul (Morus sp.) või ananassil (Ananas sp.). Kui perikarp on lihakas, räägitakse lihakviljadest, kui aga kuiv, siis kuivviljadest. Harvad pole juhtumid, kui algselt lihakas vili areneb kuivviljaks. Ühtset viljade klassifikatsiooni pole, seetõttu piirdume vaid üksikute viljatüüpide anatoomia lähema vaatlemisega.
Lihakviljad moodustuvad nii ühe- kui mitmepesalisest apo- või sünkarpsest sigimikust. Perikarp koosneb kas ainult sigimikuseinast või lisanduvad sellele ka sigimikuvälised koed. Lihakaks võib muutuda kogu perikarp või ainult osa sellest.
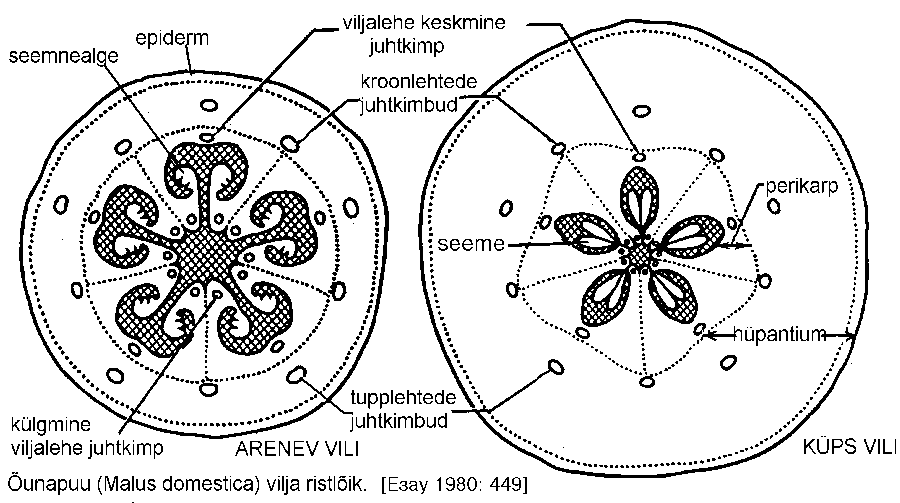
Õunvili (malum) on iseloomulik roosõieliste (Rosaceae) sugukonna õunapuuliste alamsugukonnale Pomoideae, näiteks perekondades õunapuu (Malus), pirnipuu (Pyrus), pihlakas (Sorbus), küdoonia (Cydonia), viirpuu (Crataegus). Õunvili areneb alumisest sigimikust, kusjuures enamik lihakast osast moodustub laienenud õiepõhja servadest -- hüpantiumist.
Õunvilja viiepesalise "südame" pärgamendisarnased seinad vastavad viljalehtede seintele. Väljast on noor õunvili kaetud kutiniseerunud epidermiga, esinevad ka kloroplastid, õhulõhed ja trihhoomid. Edasises arengus asenduvad õhulõhed lõvedega, kloroplastid kromoplastidega, epiderm aga õhukese peridermiga, moodustades õuna "naha".
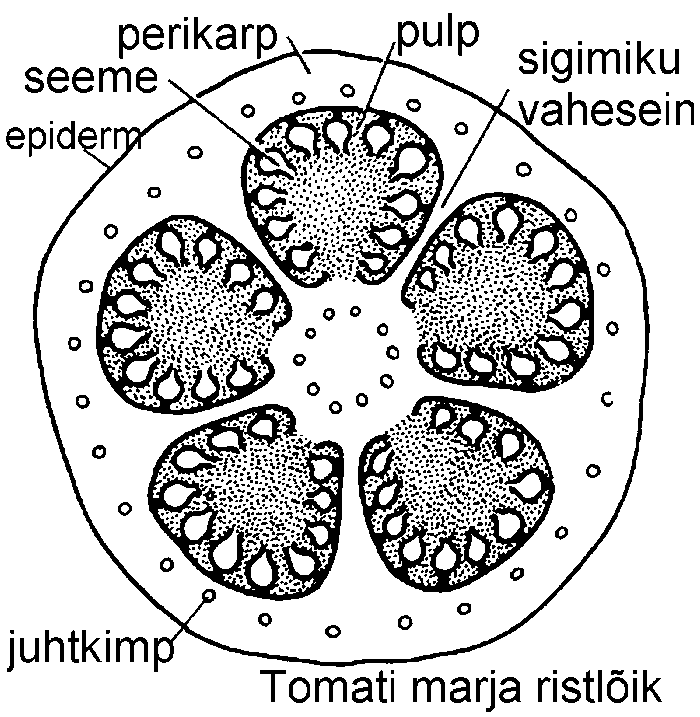
Mari (bacca) esineb näiteks maavitsalistel (Solanaceae). Tomati (Lycopersicon esculentum) kultuursortidel on mari moodustunud rohkem kui kahest viljalehest, kuigi maavitsalistele on iseloomulik kahe viljalehe esinemine. Kultuursortide viljalehtede rohkust põhjustab mitme õie kokkukasvamine. Tomati viljade valmimisel asendub klorofüll peamiselt karotinoid lükopiiniga, mis annab küpsetele viljadele kollase või punase värvi. Tomati marja mahlakas, poolvedel osa -- pulp on tekkinud peamiselt platsentast.
 Luuvili (drupa) esineb roosõieliste (Rosaceae)
ploomipuuliste (Prunoideae) alamsugukonnas,
näiteks perekondades ploomipuu (Prunus),
kirsipuu (Cerasus), toomingas (Padus),
aprikoosipuu (Armeniaca) ja virsik (Persica).
Ploomide luuvili areneb ühest viljalehest, viljalehe
servade liitekoht on vilja pinnal näha vaona.
Luuvilja perikarp on hõlpsasti jaotatav õhukeseks
eksokarbiks (koosneb kutiniseerunud, vahakihiga
kaetud epidermist ja subepidermaalsest
kollenhüümist), mahlaseks mesokarbiks ning
kivistunud endokarbiks ("luu"). Endo- ja
mesokarbis leidub juhtkimpe. Vaarika (Rubus
idaeus) koguluuvili koosneb üksikutest
osaluuviljadest.
Luuvili (drupa) esineb roosõieliste (Rosaceae)
ploomipuuliste (Prunoideae) alamsugukonnas,
näiteks perekondades ploomipuu (Prunus),
kirsipuu (Cerasus), toomingas (Padus),
aprikoosipuu (Armeniaca) ja virsik (Persica).
Ploomide luuvili areneb ühest viljalehest, viljalehe
servade liitekoht on vilja pinnal näha vaona.
Luuvilja perikarp on hõlpsasti jaotatav õhukeseks
eksokarbiks (koosneb kutiniseerunud, vahakihiga
kaetud epidermist ja subepidermaalsest
kollenhüümist), mahlaseks mesokarbiks ning
kivistunud endokarbiks ("luu"). Endo- ja
mesokarbis leidub juhtkimpe. Vaarika (Rubus
idaeus) koguluuvili koosneb üksikutest
osaluuviljadest.
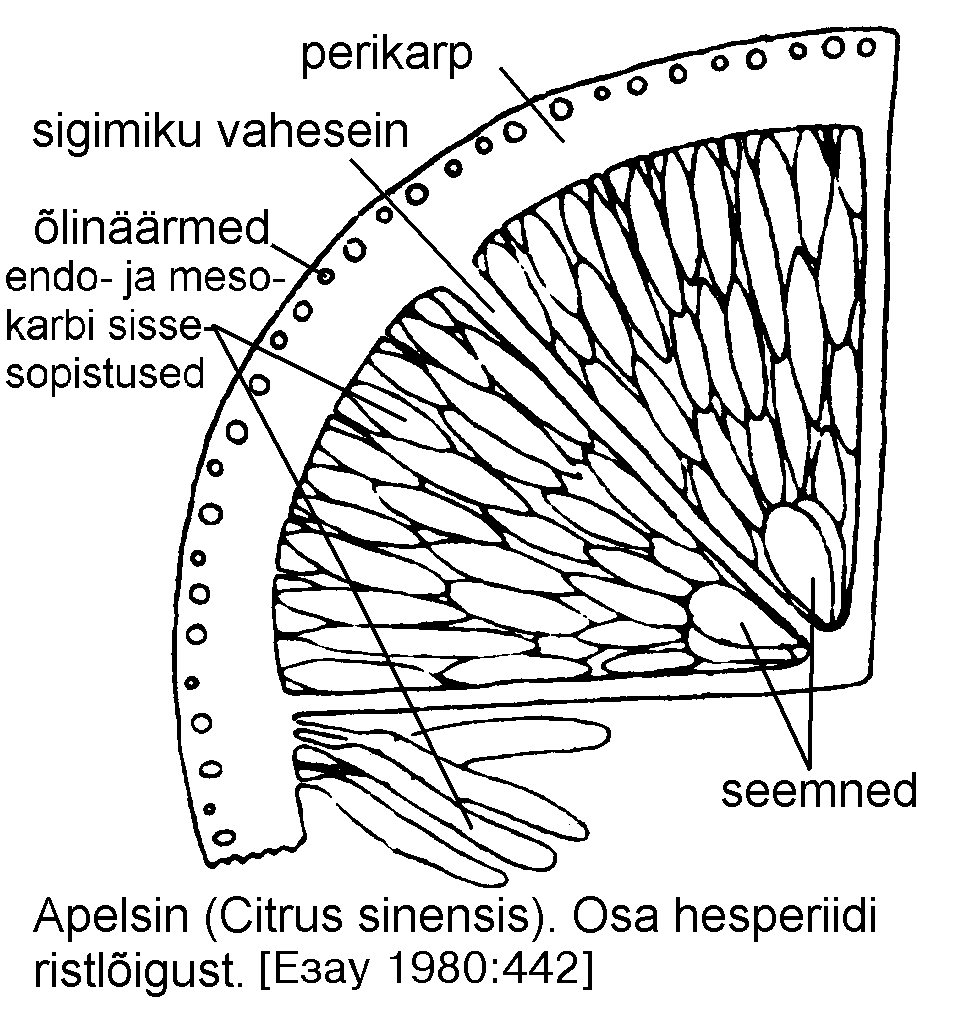
Hesperiid (hesperidium), nimetatud ka pomerantsiks (aurantium), esineb ruudiliste (Rutaceae) sugukonnas tsitruse (Citrus) perekonnas. Vili areneb umbes kümnest viljalehest. Esineb tsentraalne marginaalne platsentatsioon. Vilja ümbritsev kollane kude -- eksokarp ehk flavedo koosneb kutiniseerunud epidermist ja tihedast subepidermaalsest põhikoest, milles esineb õlikäike ja kristalle. Valge mesokarp ehk albedo on suurte rakuvaheruumidega, haruliste jätketega rakkudest moodustunud aerenhüüm. Endokarp koosneb epiteelist ja mõnest tihedast põhikoekihist. Sigimikupesade vahel asetsevad kelmed kujutavad endast endo- ja mesokarbi jätkeid. Valminud vilja "koor" koosneb ekso- ja mesokarbist. Endokarbis paiknevad mahlaga täitunud kotikesed.
Kuivviljad jaotatakse kaheks põhitüübiks: avaviljad, kus seemned ei ole viljaga kokkukasvanud, seemnete väljapääsemiseks tekivad viljakesta vastavad avad ning sulgviljad, mille seemne- ja viljakestad on kokku kasvanud, nii, et seeme viljakestadest ei eraldu.
Sagedamini esinevateks avaviljadeks on kukkur, kõder, kaun, kupar ja karp, mis võivad avaneda kas
septitsiidselt, lõhe tekib piki viljalehtede kokkukasvamise piiri;
lokulitsiidselt, viljalehed lõhenevad selgmiselt;
rõngasjalt, ava läbib ristisuunaliselt kõiki viljalehti või
poorjalt, kui vilja avanemisel tekib erisugune lokaalne ava.
Kukkur (folliculus) on ühepesaline, apokarpsest sigimikust tekkinud paljuseemneline avavili, lõhe tekib piki viljalehe serva. Kui ühest õiest tekib kukkurvilju rohkem kui kaks, moodustavad need kogukukru (multifolliculus). Kukkurviljad esinevad sugukondades tulikalised (Ranunculaceae), roosõielised (Rosaceae), magnoolialised (Magnoliaceae) ning paksulehelised (Crassulaceae).
Kõder (siliqua) ja kõdrake (silicula) on kahepesalised, kahest viljalehest moodustunud viljad. Seemned kinnituvad sigimikukambri seinale. Kõdra pikkus ületab laiuse vähemalt neli korda, kõdrakese laiuse suhe pikkusesse on väiksem. Esinevad ristõieliste (Brassicaceae) sugukonnas.
Kaun (legumen) areneb ühest viljalehest ning sarnaneb kukrule, kuid avaneb kahelt poolt -- piki viljalehe liitekohta ning mööda viljalehe selgmist juhtkimpu. Kauna ja kukru vahel pole sageli võimalik ranget vahet teha, esinevad üleminekud.
Kupar (capsula) on ühe või mitmepesaline, vähemalt kolmest viljalehest kujunenud avavili. Kupar avaneb enamasti kas septitsiidselt või lokulitsiidselt. Kui tekkiv ava on rõngasjas, nimetatakse sellist vilja karbiks.
Sulgviljadest on tavalisemad pähklike, pähkel, tõru, seemnis, teris ning jaguvili.
Pähklike (nucula) moodustub ühest viljalehest ja on enamasti ühe-, harvem aga kahe- või rohkemaseemneline. Sageli esineb kogupähklike (multinucula). Maasikas (fragum) on kogupähklikese erijuht, siin on lihakale, tugevasti arenenud õiepõhjale kinnitunud hulgaliselt pisikesi pähklikesi. Maasikvili esineb ainult perekonnas maasikas (Fragaria). Kui pähklikesed on ümbritsetud õiepõhjast (täpsemalt hüpantiumist) tekkinud lihakatest kudedest, kujuneb tõrsik ehk tsünarrood (cynarrhodion), mis esineb perekonnas kibuvits (Rosa).
Pähkli (nux) tunneb ära tugeva puitunud või nahkja viljakesta järgi. Pähkel areneb sünkarpsest kahe- või kolmepesalisest sigimikust, millest fertiilne on vaid üks sigimikupesa. Pähklis asub enamasti üks, harva kaks seemet. Pähkel on iseloomulik kaseliste (Betulaceae), jalakaliste (Ulmaceae), pöögiliste (Fagaceae) ja kirburohuliste (Polygonaceae) sugukondadele. Pähkli külge kasvanud viljalehed võivad moodustada tiivad. Selliseid tiivulisi pähkleid esineb perekondades kask (Betula), lepp (Alnus) jmt.
Tõru (glans) moodustub kolmepesalisest sigimikust, milles kaks pesa jäävad viljatuks. Tõru üht otsa katab lüdi (cupula), mis areneb tsümoidse õisiku steriilsetest, viljale ligistunud harudest. Tõru esineb näiteks tammel (Quercus).
Seemnise (achenium) vilja- ja seemnekestad on tihedasti kokku kasvanud. Seemnis on iseloomulik näiteks korvõieliste (Asteraceae), palderjaniliste (Valerianaceae), kanepiliste (Cannabaceae), uniohakaliste (Dipsacaceae), nõgeseliste (Urticaceae) ja lõikheinaliste (Cyperaceae) sugukondadele. Tarnade (Carex) seemnist katab õiekattest moodustunud põisik (utriculus).
Teris (caryopsis) areneb parakarpsest, kolme viljalehega sigimikust. Terise ainukest seemet ümbritseb õhuke perikarp. Vilja katavad õiekatte -- sõkalde ja liblede jäänused. Teris on iseloomulik kõrreliste (Poaceae) sugukonnale. Terise siseehitust tutvustab pikemalt töö nr. 93.
Jaguvili ehk skisokarp koosneb kahest karpofooriga ühendatud osaviljast ehk merikarbist. Osavili sarnaneb siseehituselt seemnisele. Jaguvili esineb näiteks sugukonnas sarikalised (Apiaceae).
Sõltuvalt sellest, kas seeme (semen) areneb soomuste kaenlas (squama seminifera) või kinnises emakas, jagatakse seemnetaimed kahte hõimkonda -- paljasseemnetaimed (Pinophyta ehk Gymnospermae) ning katteseemnetaimed (Magnoliophyta ehk Angiospermae). Allpool vaatleme lähemalt katte-seemnetaimede seemneid.
Seemnete väliskuju ja suurus on väga varieeruvad. Esineb mõnekümne sentimeetrise läbimõõduga (palmilised, Arecaceae) kui ka mikroskoopilisi, tuulega levivaid tolmpeeneid seemneid (käpalised, Orchidaceae; võõrikulised, Loranthaceae). Kujult on seemned sagedamini ümarad, piklikud, kerajad, munajad või ovaalsed. Seemnete kuju sõltub sageli vilja kujust ning seemnete hulgast ühes viljas.
Kaheliviljastumise tagajärjel tekivad seemnealgmes idu (uus sporofaas) ning varuained uue taime varaseks kasvuks. Idu areneb lootekotist. Toitekoeks võib olla endosperm (tekib lootekoti teistuuma viljastamisel) ja/või perisperm (tekib nutsellist, lähemalt varuainete paigutusest allpool). Integumendid moodustavad seemnekesta. Mikropüül harilikult kaob või täitub põhikoega. Seemnealgme jalg võib jääda seemne küljele õmblusena (hilum).
Idu (embryo) on seemne kõige olulisem osa. Idu koosneb idulehtedest (cotyledones), hüpokotüülist (hypocotylus), idupungast (plumula) ja idujuurest (radicula). Okaspuudel esineb idu basaalses osas veel ühe- või mitmerakuline moodustis -- suspensor (suspensor), mille abil idu kinnitub lootekoti külge ja saab emataimelt toitaineid.
Idulehtede arvust sõltuvalt eristatakse kaht katteseemnetaimede klassi -- üheidulehelised (Liliopsida ehk Monocotyledones) ning kaheidulehelised (Magnoliopsida ehk Dicotyledones). Üheidulehelisuse teke on katteseemnetaimede evolutsioonilise morfoloogia üheks põhiküsimuseks, millele on pühendatud hulgaliselt spetsiaalseid uurimusi. Paljasseemnetaimedel (Pinophyta) on idulehti ühest (palmlehikulised -- Cycadaceae) kaheksateistkümneni (männilised -- Pinaceae).
 Idu võib olla nii suur, et täidab kogu seemne
(joonis), näiteks tammel (Quercus) või
luigelillel (Butomus), aga ta võib olla ka
väga väike, näiteks kuslapuul (Lonicera) või
piipheinal (Luzula) või koguni mikroskoopiline, nagu käpalistel (Orchidaceae).
Idu võib olla nii suur, et täidab kogu seemne
(joonis), näiteks tammel (Quercus) või
luigelillel (Butomus), aga ta võib olla ka
väga väike, näiteks kuslapuul (Lonicera) või
piipheinal (Luzula) või koguni mikroskoopiline, nagu käpalistel (Orchidaceae).
Idu on seemnes paigutunud kas selle alusele, (näiteks porgandil -- Daucus), tippu (näiteks õlipuul -- Olea), seemne keskele (okaspuudel -- Pinopsida; kamaral -- Odontites) või seemne küljele (tatralistel -- Polygonaceae). Idu on sagedamini sirge, näiteks jänesekapsal (Oxalis), kuid esineb ka käändunud, rõngasjaid või spiraliseerunud idusid.
Seemne üheks oluliseks funktsiooniks on areneva idu varustamine toitainetega. Seemne esinemine loob seemnetaimedele olulise eelise sõnajalgtaimede ees: seemned sisaldavad varuaineid, eosed aga mitte. Seega on eoste kasvamaminek ja noore taime ellujäämine sõnajalgtaimede puhul vähem tõenäoline. Seemnete varuained võivad paikneda:
1) tugevasti arenenud hüpokotüülis, näiteks sugukonnas penikeelelised (Potamogetonaceae);
2) idulehtedes, näiteks sugukondades kõrvitsalised (Cucurbitaceae), liblikõielised (Fabaceae), korvõielised (Asteraceae), pöögilised (Fagaceae);
3) endospermis (endospermium), näiteks sugukondades kanarbikulised (Ericaceae), kõrrelised (Poaceae), liilialised (Liliaceae), amarüllilised (Amaryllidaceae) jt. Endospermi olemasolu või puudumise järgi ei saa otsustada sugukonna primitiivsuse üle, sest see esineb nii primitiivsetes kui ka kõrgeltarenenud sugukondades. Okaspuudel tekib endosperm enne viljastumist, rakud on haploidsed erinevalt katteseemnetaimede triploidsest endospermist;
4) perispermis (perispermium) näiteks sugukondades nelgilised (Caryophyllaceae), kaktuselised (Cactaceae), pipralised (Piperaceae), agaavilised (Agavaceae), kirburohulised (Polygonaceae) jt.;
5) endospermis ja perispermis, endosperm on sel juhul enamasti perispermist vähem arenenud, näiteks sugukondades hundinuialised (Typhaceae), kannalised (Cannaceae);
6) kalatsospermis (chalazospermium), kui kalatsa on arenenud varuainete säilituskohaks. See nähtus on teada vaid perekonnas Cyanastrum.
Üldreeglina on varuaineid seda vähem, mida paremini on idu arenenud. Endosperm ja perisperm on erinevates sugukondades väga mitmesuguse konsistentsi ja struktuuriga. Konsistents sõltub ka seemne füsioloogilisest seisundist -- kas seeme on puhkeolekus või hakkab juba idanema.
Seemnekest (spermoderm) on enamasti tekkinud integumentidest, mõnikord võtavad seemnekesta moodustamisest osa ka nutsell ja kalatsa. Üksikute sugukondade esindajatel võib seemnekest puududa, näiteks võõrikulised (Loranthaceae). Enamasti on seemnekest mitmekihiline (roosõielised Rosaceae, ruudilised Rutaceae jt.), kuid võib koosneda ka ühest rakukihist (käpalised Orchidaceae, kanarbikulised Ericaceae jt.). Seemnekesta pealispind võib olla väga erineva struktuuri, konsistentsi ja värvusega ning võib kanda ka mitmesuguseid lisandeid -- olla karvane, väljakasvetega jne.
Seemnetel esineb mitmesuguseid jätkeid, mis kujutavad endast kas seemnekesta väljakasveid, seemnenaba või seemnealgme jala tekiseid. Sama päritoluga on ka tiivad ja arillus ehk seemnerüü (arillus). Tiibu võib seemnel olla üks, kaks või mitu. Arillus on enamasti lihakas, sisaldab rasvu, suhkruid, tärklist jt. toitaineid, mistõttu sellest toituvad meelsasti mitmesugused loomad, aidates seeläbi kaasa seemnete levimisele. Arillus võib seemne üleni katta, võib aga olla ka nii väike, et vaevalt nähtav.
Seemned on enamasti pruuniks või mustaks värvunud, harva esineb valgeid, punaseid, kollaseid ja rohelisi seemneid. Kõik loetletud värvused on seotud üleminekutega. Seemnele lisab värvi ka mitmeti värvunud arillus. Seemne väliskuju on enamasti seotud seemne levimisviisiga.
Seemnete põhiline bioloogiline funktsioon on anda algus uuele taimele, kaitsta idu ebasoodsate tingimuste eest ja kindlustada idanevale taimele toitainevarud. Tähtis on ka levila laiendamise funktsioon seemnete ja viljade kauglevi teel.
Kõiki taimeosi, millest võib areneda uus taim, nimetatakse levisteks ehk diaspoorideks. Nendeks võivad olla seemned, viljad, vilikonnad, harvem taime suuremad osad koos viljadega, samuti taime vegetatiivsed osad koos pungadega. Diaspooride levimisviisidest tuleks nimetada eelkõige järgmisi:
1) autohooria ehk iselevi, kohastumused diaspooride levitamiseks viljumiskohast kaugemale puuduvad, seemned kukuvad maha emataime lähedusse, näiteks perekondades nurmenukk (Primula) või tulp (Tulipa);
2) anemohooria ehk tuullevi korral esinevad mitmesugused kohastumused levimiseks tuule abil, näiteks lendkarvad pajul (Salix), tiivad upsujuurel (Veratrum), ääris liilialistel (Liliaceae), või on seemned sedavõrd väiksed, nii et tuul kannab neid ka ilma abivahenditeta, näiteks sookailul (Rhododendron tomentosum);
3) hüdrohooria ehk vesilevi -- diaspoore kannab edasi vesi;
4) zoohooria ehk loomlevi -- diaspoore levitavad mitmesugused loomad. Sõltuvalt loomarühmast eristatakse
a) ornitohooriat ehk lindlevi, kui diaspoore levitavad linnud,
b) mürmekohooria ehk sipelglevi, diaspoore levitavad sipelgad, kes toituvad seemnete küljes olevatest õlikehadest -- elaiosoomidest.
Epizoohooria korral on seemned haakjate väljakasvetega või kleepuvad, diaspoorid levivad looma välispinnale kinnitunult. Endozoohoorsed seemned on enamasti ümbritsetud mahlakate söödavate viljadega. Seemned on paksukestalised, mistõttu need läbivad looma seedekulgla idanevust kaotamata.
1. Õpikuid ja monograafiaid:
Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K. Watson, J. D. 1989. Molecular Biology of the Cell. 2nd ed. Garland Publ., New York/London.
Bell A. D. 1991. Plant Form. Oxford University Press, Oxford. 341 p.
Bold, H. C., Alexopoulus, C. J., Delevorvas, T. 1987. Morphology of plants and fungi. 5th ed. Harper & Row, New York. X+912 pp.
Eames, A. J. 1961. Morphology on the Angiosperms. McGraw-Hill Book Company, New York/Toronto/London. [Sama raamat on ilmunud ka vene keeles: Èìñ, À. 1961. Ìîðôîëîãèÿ öâåòêîâûõ ðàñòåíèé. Ìèð, Ìîñêâà. 497 ñ.]
Eames, A. J., McDawels, L. M. 1951. An Introduction to Plant Anatomy. N.Y. McGraw-Hill Book Company, London.
Esau, K. 1965. Plant anatomy. 2. ed. John Wiley & Sons, Inc. New York/London/Sydney.
Esau, K. 1977. Anatomy of Seed Plants. 2nd ed. John Wiley and Sons, New York/Santa Barbara/London/Sydney/Toronto. [Sama raamat on ilmunud ka vene keeles: Ýçàó, Ê. 1980. Àíàòîìèÿ ñåìåííûõ ðàñòåíèé. ò. 1-2. Ìèð, Ìîñêâà. 558 ñ.]
Fahn, A. 1990. Plant anatomy. 4th ed. (1st 1967), Pergamon Press, Oxford. xi+588 p.
Haberlandt, G. 1918. Physiologische Pflanzenanatomie. 5., neubearbeitete und vermehrte Aufl. Wilhelm Engelmann, Leipzig. 670 S.
Kalda, A., Kukk, E., Masing, V., Trass, H., Vaga, A. 1965. Botaanika. Õpik kõrgematele koolidele. I. osa: Taimetsütoloogia, -anatoomia ja -morfoloogia. Koostanud H. Trass. Valgus, Tln., 432 lk.
Kaussmann, B. 1963. Pflanzenanatomie unter besonderer Berücksichtigung der Kultur- und Nutzpflanzen. Gustav Fischer Verlag, Jena. 624 S.
Kaussmann, B., Schiewer, U. 1989. Funktionelle Morphologie und Anatomie der Pflanzen. Gustav Fischer Verlag, Jena. 465 S.
Lehrbuch der Botanik für Hochschulen. 1991. E. Strasburger, F. Noll, H. Schenck, A. F. W. Schimper, neubearbeitet von P. Sitte, H. Ziegler, F. Ehrendorfer, A. Bersinsky. 34. Aufl. Gustav Fischer Verlag, Stuttgart-Jena-New York. 1030 S. [Teos on välja antud paljude kordustrükkidena saksa, inglise, hispaania, poola, serbo-horvaadi, tðehhi, vene, jaapani ja itaalia keeles. Eesti keeles ilmus raamatu esimene osa 1926. a. Tartus (tõlgiti 16. trüki järgi)].
Lippmaa, T. 1940. Üldine botaanika. Tartu, Akadeemilise Kooperatiivi Kirjastus. 358 lk.
Nadakavukaren, M.; McCracken, D. 1985. Botany. An Introduction to Plant Biology. West Publishing Company. Saint Paul-New York-Los Angeles-San Francisco. 591 p.
Raven, P. H., Evert, R. F, Eichorn, S. E. Biology of plants. 4th ed. Worth publishers, Inc. [See raamat on hiljuti ilmunud ka vene keeles: Ðåéâí, Ï., Ýâåðò, Ð., Àéõîðí, Ñ. 1990. Ñîâðåìåííàÿ áîòàíèêà. Â 2-õ òîìàõ. Ìèð, Ìîñêâà. ò. 1. 348 ñ.]
Rudall, P. 1992. Anatomy of flowering plants: An introduction to structure and development. 2nd ed. Cambridge University Press, Cambridge. ix+110 p.
Weier, T. E, Stocking, C. R. Barbour, M. G. 1974. Botany. An Introduction to Plant Biology. 5th Ed. J. Wiley & Sons, New York-Chichester-Brisbane-Toronto. 693 p.
Àòàáåêîâà, À. È., Óñòèíîâà, Å. Ë.1987. Öèòîëîãèÿ ðàñòåíèé. Èçä. 4., ïåðåðàá. è äîïîë. Àãðîïðîìèçäàò, Ìîñêâà. 246 ñ.
Áàííèêîâà, Â. Í., Õâåäûíèũ, Î. À. 1982. Îñíîâû ýìáðèîëîãèè ðàñòåíèé. Íàóêîâà Äóìêà, Êèåâ. 164 ñ.
Âàñèëüåâ, À. Å., Âîðîíèí, È. Ñ., Åëåíåâñêèé, À. Ã., Ñåðåáðÿêîâà, Ò. È., Øîðèíà, Í. È. 1988. Áîòàíèêà. Ìîðôîëîãèÿ è àíàòîìèÿ ðàñòåíèé. 2-å èçä., ïåðåðàá. Ïðîñâåùåíèå, Ìîñêâà. 480 ñ.
Æóêîâñêèé, Ï. Ì. 1982. Áîòàíèêà. Èçä. 5., ïåðåðàá. è äîïîë. Êîëîñ, Ìîñêâà. 624 ñ.
Ñàâũåíêî, Ì. È. 1973. Ìîðôîëîãèÿà ñåìÿïîũêè ïîêðûòîñåìåííûõ ðàñòåíèè. Íàóêà, Ëåíèíãðàä. 110 ñ. [Põhjalik monograafia seemnealgme ehitusest]
Òóòàþê, Â. Õ. 1980. Àíàòîìèÿ è ìîðôîëîãèÿ ðàñòåíèé. Èçä. 2., ïåðåðàá. è äîïîë. Âûñøàÿ øêîëà, Ìîñêâà. 318 ñ.
2. Praktikumide juhendeid ning muid abimaterjale:
Braune, W., Leman, A., Taubert, H. 1991. Pflanzenanatomisches Praktikum I. Zur Einführung in die Anatomie der Vegetationsorgane der Samenpflanzen. 6. Aufl. Gustav Fischer Verlag, Jena. 283 S.
Braune, W., Leman, A., Taubert, H. 1976. Praktikum zur Morphologie und Entwicklungsgeschichte der Pflanzen. VEB Gustav Fischer Verlag, Jena. 448 S.
Cresti, M., Blackmore, S., van Went, J. L. 1992. Atlas of Sexual Reproduction in Flowering Plants. Springer-Verlag, Berlin. xi+249 p.
Foster, A. S. 1949. Practical Plant Anatomy. D. van Nostrand Company. Toronto, New York, London.
Järvekülg, L., Riispere, A. 1962. Kromosoomide arvu määramisest taimedel. Eesti Loodus nr. 5, lk. 299-301.
Komarov, V. L. 1946. Praktiline taimeanatoomia. Tlk. J. Talts. Teaduslik Kirjandus, Tartu. 360 lk.
Kukk, E., Lasting, V. 1959. Kuidas käsitseda mikroskoopi. -- Eesti Loodus nr. 2, lk. 119-122.
Lacey A. J. (ed.) 1991. Light microscopy in biology: a practical approach. IRL Press at Oxford University Press, Oxford. xviii+329 p.
Mader, S. S. 1991. Inquiry into life. Laboratory Manual. 4th Ed. W. C. Brown Publ. Dubuqure-Iowa. 368 p.
Meyer, A. 1915. Erstes mikroskopisches Praktikum. Eine Einführung in den Gebrauch des Mikroskopes und in die Anatomie der höheren Pflanzen. Zum Gebrauche in den botanischen Laboratorien und zum Selbstunterrichte. 3., vervollständigte Aufl. Verlag von Gustav Fischer, Jena. 255 S.
Natho, G., Müller, C., Schmidt, H. 1990. Morphologie und Systematik der Pflanzen. 1-2 Teil. G. Fischer Verlag. 850 S.
Neerut, H. 1974. Taimeanatoomia praktikum. 2., täiend. ja ümbertöötatud tr. Tartu, EPA, 76 lk.
Nultsch, W., Grahle, A. 1988. Mikroskopisch-Botanisches Praktikum für Anfänger. 8., überarbeitete Aufl. Georg Thieme Verlag, Stuttgart/New York. 207 S.
Paaver, U., Raal, A. 1995. Droogide mikroskoopia. Metoodiline juhend arstiteaduskonna farmaatsiaosakonna üliõpilastele farmakognoosia praktikumideks. Tartu Ülikool, Tartu. 149 lk.
Pyykkö, M. 1983. 52549 Puuanatomian kurssi. Biologian koulutusohjelman vaihtoehtoinen ainepintojakso. Helsingin yliopiston kasvitieteen laitoksem monisteita 84. Helsnki, 71 p.
Port, J. 1936. Praktiline taimeanatoomia ja -füsioloogia. K./Ü. Loodus, Tartu. 106 lk.
Ratt, A., Reitan, V. 1969. Kultuurtaimede ja umbrohtude seemned. Valgus, Tallinn. 260 lk. + 24 tahvlit.
Saar, A. 1962. Botaaniliste püsipreparaatide valmistamise metoodika. TRÜ geneetika ja darvinismi kateeder. Tartu. 20 lk.
Stamm, J., Toomingas, A. 1936. Farmakoanatoomia. Akadeemilise Kooperatiivi Kirjastus, Tartu. viii+146 lk.
Strasburger, E., Koernicke, M. 1921. Das Kleine botanische Praktikum für Anfänger. 9., verbesserte Aufl. Gustav Fischer, Jena. 272 S.
Strasburger, E., Koernicke, M. 1923. Das botanische Praktikum. 7. Aufl. Gustav Fischer, Jena. 883 S.
Veermets, K. 1962. Puidu määraja makro- ja mikroskoopiliste tunnuste järgi. 2. tr. Eesti Riiklik Kirjastus, Tallinn. 128 lk.
Áàâòóòî, Ã. À. 1985. Ëàáîðàòîðíûé ïðàêòèêóì ïî àíàòîìèè è ìîðôîëîãèè ðàñòåíèé. Âûøýéøàÿ øêîëà, Ìèíñê. 352 ñ.
Áàðûêèíà, Ð. Í., Êîñòðèêîâà, Ë. È., Êîũåìàðîâà, È. Ï., Ëîòîâà, Ë. È., Òðàíêîâñêèé, Ä. À., Ũèñòÿàêîâà, Î. È. 1979. Ïðàêòèêóì ïî àíàòîìèè ðàñòåíèé. 3-å èçä., ïåðåðàá. è äîïîë. Âûñøàÿ øêîëà, Ìîñêâà. 224 ñ.
Áåðåçîâñêàÿ, Ò. Ï., Äîùèíñêàÿ, Í. Â., Ñåðûõ, Å. À. 1978. Ìåòîäû ìèêðîñêîïèũåñêîãî àíàëèçà áîòàíèũåñêèõ îáúåêòîâ (óũåáíî-ìåòîäèũåñêîå ïîñîáèå ïî áîòàíèêå). Òîìñê. 115 ñ.
Äæàïàðèäçå, Ë. È. 1953. Ïðàêòèêóì ïî ìèêðîñêîïèũåñêîé õèìèè ðàñòåíèé. Ñîâåòñêàÿ íàóêà, Ìîñêâà. 152 ñ.
Êèñåëåâà, Í. Ñ., Øåëóõèí, Í. Â. 1969. Àòëàñ ïî àíàòîìèè ðàñòåíèé, Âûøýéøàÿ øêîëà, Ìèíñê. 288 ñ.
Ëîòîâà, Ë. È., Òèìîíèí, À. K. 1990. Aíàòîìèÿ ñòåáëåé è âòîðèũíûõ ïðîâîäÿùèõ òêàíåé äðåâåñíûõ ðàñòåíèé. Èçä. Ìîñêîâñêîãî Óíèâåðñèòåòà, Ìîñêâà. 58 ñ.
Íàóìîâ, Í. À., Êîçëîâ, Â. Å. 1954. Îñíîâû áîòàíèũåñêîé ìèêðîòåõíèêè. Ñîâåòñêàÿ íàóêà, Ìîñêâà. 312 ñ.
Íèêèòèí, À. À., Ïàíêîâà, È. À. 1982. Àíàòîìèũåñêèé àòëàñ ïîëåçíûõ è íåêîòîðûõ ÿäîâèòûõ ðàñòåíèé. Íàóêà, Ëåíèíãðàä. 767 ñ.
Ïàóøåâà, Ç. Ï. 1988. Ïðàêòèêóì ïî öèòîëîãèè ðàñòåíèé. Èçä. 4., ïåðåðàá. è äîïîë. Àãðîïðîìèçäàò, Ìîñêâà. 272 ñ.
Ïðîçèíà, Ì. Í. 1960. Áîòàíèũåñêàÿ ìèêðîòåõíèêà. Âûñøàÿ øêîëà, Ìîñêâà. 207 ñ.
Òåðïèëî, Í. È. 1961. Àíàòîìèũåñêèé àòëàñ ëåêàðñòâåííûõ ðàñòåíèé. 2., ïåðåðàá. è äîïîë. èçä. Ãîñóäàðñòâåííîå ìåäèöèíñêîå èçä. ÓÑÑÐ. Êèåâ. 363 ñ.
Õðæàíîâñêèé, Â. Ã., Ïîíîìàðåíêî, Ñ. Ô. 1989. Ïðàêòèêóì ïî êóðñó îáùåé áîòàíèêè. Èçä. 2., ïåðåðàá. è äîïîë. Àãðîïðîìèçäàò, Ìîñêâà. 416 ñ.