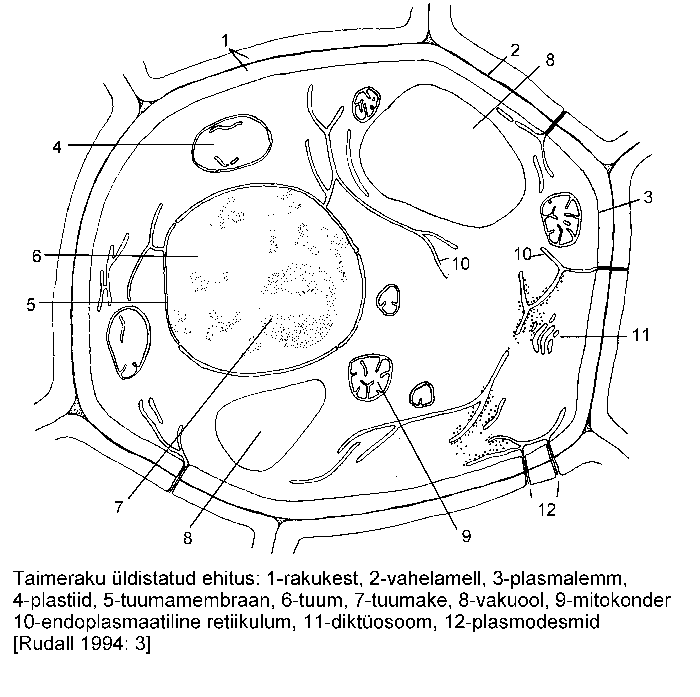
Soontaimed, nagu kõik teisedki hulkraksed eukarüoodid, koosnevad hästi eristuvatest üksustest -- rakkudest (joonis), mis üheskoos moodustavad funktsionaalse terviku. Rakkude kuju, suurus, struktuur ja funktsioonid on väga mitmekesised.
Kõrgemate taimede rakke on kuju järgi võimalik jagada kaheks põhitüübiks -- parenhüümsed ehk isodiameetrilised rakud, mille läbimõõt on igas suunas enam-vähem võrdne ning prosenhüümsed rakud, mille pikkus ületab tunduvalt laiuse. Rakkude läbimõõt on enamasti 10-100 µm, pikkus võib kiudtaimede prosenhüümsetel rakkudel ulatuda 25 cm-ni (bömeeria ehk ramjee, Boehmeria nivea). Rakkude suurus on koele iseloomulik tunnus, mis ei sõltu taime suurusest (suure taime rakud pole väiksema taime rakkudest suuremad).
Tinglikult on taimerakk jaotatav elusosaks -- protoplastiks ning eluta osaks (rakukest, varuained, eritusained jms.). Protoplastis eristatakse membraan-, fibrillaar- ja makromolekulaarseid (kromosoomid, ribosoomid) struktuure. Läbipaistev vedelik, mis täidab raku sisu ning milles paiknevad raku organellid, on tsütosool (nimetatud ka põhiaineks ehk maatriksiks). Tsütoplasmaks nimetatakse tsütosooli koos organellidega, kuid ilma tuumata. Tsütoplasma tihedamat väliskihti, kus paikneb tihe aktiinifilamentide võrgustik, nimetatakse kortikaalseks tsütoplasmaks.
Taimerakkude kaks põhilist ehituslikku ja talitluslikku erinevust, võrrelduna loomarakuga, on tingitud mitmesuguste plastiidide ja jäiga rakukesta olemasolust. Samas tuleb aga märkida, et rakkude põhilised eluavaldused -- DNA replikatsioon, samuti valgusüntees ning membraanide ehitus on nii taime- kui ka loomarakkudel ühesugused.
Membraanstruktuurid võivad koosneda ühest või kahest membraanist. Kahe membraaniga ümbritsetud organellideks on plastiidid, mitokondrid ja tuum, kõik nimetatud organellid sisaldavad suuremal või vähemal määral DNA-d.
Plastiidides toimub fotosüntees, varuainete ümberkujundamine jt. taimele olulised protsessid. Plastiidid on pooldumisvõimelised organellid. Nende keskmine läbimõõt on 3-8 µm.
Kõik soontaimede eritüübilised plastiidid tekivad algkudedes asuvatest, muutliku kujuga läbipaistvatest kehakestest -- proplastiididest. Pimedas arenevad proplastiididest võrdlemisi vähediferentseerunud ehitusega, prolamellaarkehi ja protoklorofülli sisaldavad etioplastid. Valguse mõjul muutub etioplastide protoklorofüll klorofülliks, sünteesitakse uusi membraane, pigmente ja ensüüme ning etioplastist kujuneb kloroplast. Kloroplasti (joonis) ümbritseb kaksikmembraan. Sisemine membraan ümbritseb kloroplasti põhiainet -- stroomat. Stroomas paiknevad membraanmoodustised -- tülakoidid pole ilmselt kloroplasti sisemise membraaniga ühenduses. Tülakoidide kogumikku (mis väljanägemiselt meenutab pannkookide virna) nimetatakse graaniks. Peale selle sisaldab kloroplast lipiiditilku, ribosoome, DNA-d ja esmast ehk assimilatsioonitärklist. Tülakoidides paiknev klorofüll annab kloroplastidele rohelise värvuse.
Toored viljad on enamasti rohelised, sest need sisaldavad kloroplaste. Viljade valmimisel kaob klorofüll kloroplastidest ning mõjule pääsevad kollased või punased pigmendid -- mitmesugused karotinoidid, ning kloroplast muutub kromoplastiks (joonis). Neid esineb lisaks valminud viljadele ka õite kroonlehtedes. Kromoplastide kuju on üldiselt perekonna- või sugukonnaspetsiifiline. Taimedele ei anna värvuse mitte üksnes kloro- või kromoplastiidid, seda mõjutavad ka vakuoolides leiduvad pigmendid, näiteks antotsüaan.
Vanemas kirjanduses on eristatud ainult kloro-, kromo- ja leukoplaste, viimasteks loeti kõiki värvusetuid plastiide. Kitsas mõistes on leukoplastid vaid epidermis ja mujal leiduvad värvitud etioplastid. Säilitusfunktsiooniga leukoplaste nimetatakse kaasajal kas elaioplastideks, kui varuaineteks on õlid, amüloplastideks, kui selleks on säilitustärklis või proteinoplastideks, kui need sisaldavad varuvalku.
Mitokondreid on ühes rakus tavaliselt mitusada.
Nende kuju varieerub niitjast ellipsoidini. Väljast
on need, sarnaselt plastiididele, kaetud kahe
membraaniga. Mitokondreid täitvat põhiainet
nimetatakse maatriksiks. Oluliseks erinevuseks,
võrrelduna kloroplastiga, on sisemiselt
membraanilt mitokondri sisemusse suunduvad
mitmesugused sõrmjad moodustised või
vaheseinad -- kristad ehk harjased. Nagu
plastiidid, paljunevad ka mitokondrid jagunemise
teel, mis toimub tuuma jagunemisest sõltumatult.
Nende keskmine pikkus on 1-5 µm ning läbimõõt 0,5-1 µm. 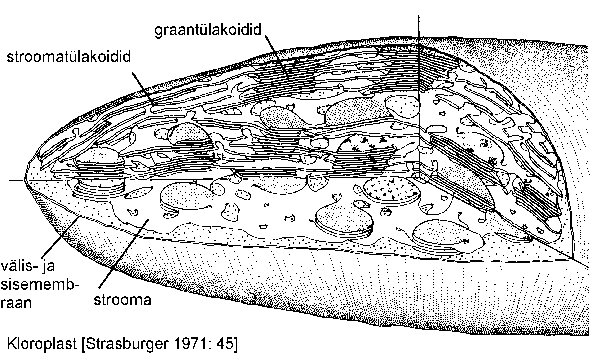
Enamasti on igas rakus üks tuum, kuid täiskasvanud elusad rakud (nt. sõeltorude rakud) võivad olla ka tuumata. Samuti esineb mitmetuumalisi rakke, näiteks õistaimede lootekott. Tuum on ümbritsetud kahe membraaniga, milles on ohtralt poore. Välimine membraan on ühenduses endoplasmaatilise retiikulumiga. Tuumaplasma (karüoplasma) põhikomponendiks on kromatiin. Sageli on tuumas üks kuni mitu tumedamat moodustist -- tuumakest (nucleolus), kus toimub ribosoomide komponentide moodustumine. Tuumade keskmine läbimõõt on 10-20 µm.
Ühekihiline membraan on iseloomulik järgmistele rakustruktuuridele: diktüosoomid, lüsosoomid, plasmalemm, tonoplast, peroksisoomid ja endoplasmaatiline retiikulum. Plasmalemm asub vahetult rakukesta all ning ümbritseb tsütoplasmat, tonoplast piirab vakuooli. Plasmalemm ja tonoplast pole valgusmikroskoobis nähtavad.
Vakuool on enamiku taimerakkude suurimaks ja silmatorkavaimaks kompartemendiks. Ühes rakus on üks või mitu vakuooli. Harilikult on vakuooliga täidetud umbes pool raku mahust, kuid sõltuvalt raku tüübist võivad need enda alla võtta 5-95%. Vakuoolid tekivad noortes algkoe rakkudes ilmselt nii diktüosoomidest kui ka endoplasmaatilisest retiikulumist. Suured vakuoolid on enamasti läbistatud tsütoplasmaväätidest. Suuri vakuoole sisaldavate rakkude tsütoplasma koos rakuorganellidega on tihedasti liibunud vastu plasmalemmi ja rakukesta.
Taimeraku vakuoolid on ehituselt ja talitluselt lähedased loomse raku lüsosoomidele. Ka vakuoolid võivad sisaldada hulgaliselt hüdrolüütilisi ensüüme. Vakooli funktsioonideks on:
1) toitainete ja jääkproduktide säilitamine;
2) teatud ainete lagundamine (sarnaselt lüsosoomiga);
3) turgori reguleerimine.
Tihtipeale on taimerakus mitu vakuooli, millest üks täidab näiteks säilitusfunktsiooni, teine aga talitleb lüsosoomina.
Diktüosoomid on tuuma lähedal asuvad lamedad
moodustised, mille põhiline ülesanne on rakukesta
komponentide süntees ja nende sekreteerimine raku
välispinnale. Loomarakkudel nimetatakse analoogilist
organelli Golgi aparaadiks. 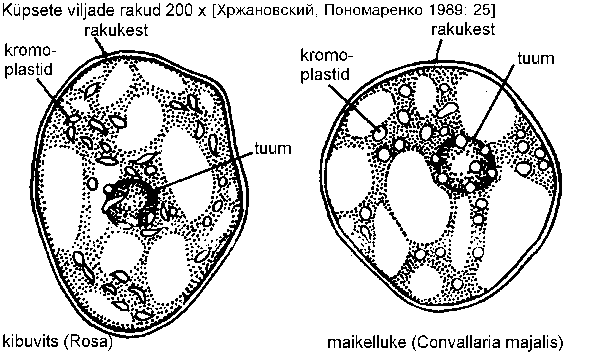
Tsütoplasmat läbib elektronmikroskoobi abil nähtav peen võrgustik -- endoplasmaatiline retiikulum (ER). See koosneb kanalikestest ning nende laienditest (tsisternidest). Kanalikesed läbivad plasmodesme -- rakukestade poorides sisalduvaid plasmaniite -- nii moodustub ühendus naaberrakkude vahel. Kohati on ER kaetud ribosoomidega, kus toimub valgusüntees (translatsioon).
Mikrokehad on väikesed (diameeter 0,5-1,5 µm), ühekordse membraaniga ümbritsetud organellid. Taimedes esineb kahesuguseid mikrokehi. Üks tüüp, peroksisoomid, on levinud leherakkudes, need osalevad fotorespiratsioonis. Teist tüüpi mikrokehi esineb idanevates seemnetes, kus need konverteerivad rasvhapped noore taime kasvuks vajalikeks suhkruteks. Kuna selline lipiididest suhkrute teke käib nn. glüoksülaat-tsükli kaudu, nimetatakse vastavaid mikrokehi glüoksüsoomideks.
Sarnaselt loomsetele rakkudele, on ka kõigis taimerakkudes olemas tsütoskelett. Selle moodustavad kolm valguliste filamentide süsteemi:
1) aktiinifilamendid e. mikrofilamendid (läbimõõt 6-8 µm), monomeeriks on globulaarne valk aktiin. Taimerakkudes esinev aktiin on molekulaarmassilt ja aminohapete järjestuselt sarnane loomarakkude aktiinile. Aktiinifilamentide osalusel leiab aset tsütoplasmaliikumine;
2) mikrotuubulid (läbimõõt 25 nm), monomeeriks on tubuliin, mis on samuti globulaarne valk. Mikrotuubulid on olulised rakkude jagunemisel, sest need moodustavad enamiku kääviniidistikust;
3) intermediaalsed filamendid (läbimõõt 10 nm), mille monomeeriks on mitmesugused fibrillaarsed valgud (tsütokeratiinid, lamiinid jt.).
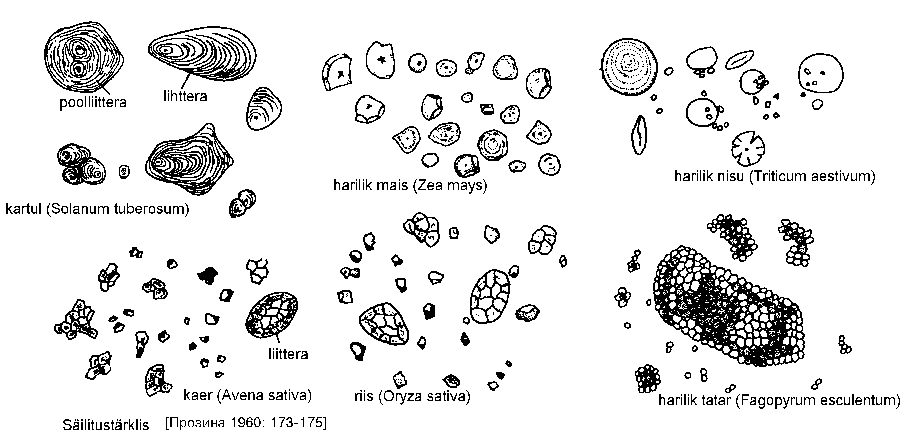 Varuainetena esinevad
taimedes säilitustärklis
(varutärklis),
aleurooniterad ning
õlitilgad. Säilitustärklis
(joonis) ladestub
amüloplastides
moodustumiskeskme
ümber. Kui
moodustumiskese ei asu amüloplasti keskel, vaid kuskil servas, ladestub tärklis
intensiivsemalt sellel küljel, kus amüloplasti strooma kiht on paksem. Nii moodustuvad
ekstsentrilised tärkliseterad (näiteks kartulil, Solanum tuberosum). Kontsentrilisi tärkliseteri
näeme kõrrelistel (Poaceae). Liittärkliseterad, mis koosnevad otsekui mitmest lihtterast,
sisaldavad mitut moodustumiskeset, kuid ühised tärklisekihid puuduvad (näiteks kaeral,
Avena sativa). Poolliitterade moodustumiskeskmed on ümbritsetud ühiste kihtidega. Öösel
ladestuvad tärklisekihid on päevastest tumedamad suurema veesisalduse tõttu. Tärkliseterad
on enamasti liigispetsiifilise kujuga, mis võimaldab mikroskoobi abil määrata jahu koostist.
Varuainetena esinevad
taimedes säilitustärklis
(varutärklis),
aleurooniterad ning
õlitilgad. Säilitustärklis
(joonis) ladestub
amüloplastides
moodustumiskeskme
ümber. Kui
moodustumiskese ei asu amüloplasti keskel, vaid kuskil servas, ladestub tärklis
intensiivsemalt sellel küljel, kus amüloplasti strooma kiht on paksem. Nii moodustuvad
ekstsentrilised tärkliseterad (näiteks kartulil, Solanum tuberosum). Kontsentrilisi tärkliseteri
näeme kõrrelistel (Poaceae). Liittärkliseterad, mis koosnevad otsekui mitmest lihtterast,
sisaldavad mitut moodustumiskeset, kuid ühised tärklisekihid puuduvad (näiteks kaeral,
Avena sativa). Poolliitterade moodustumiskeskmed on ümbritsetud ühiste kihtidega. Öösel
ladestuvad tärklisekihid on päevastest tumedamad suurema veesisalduse tõttu. Tärkliseterad
on enamasti liigispetsiifilise kujuga, mis võimaldab mikroskoobi abil määrata jahu koostist.
Taimede seemnetes kogunevat varuvalku nimetatakse aleurooniks. See esineb amorfsete aleurooniterade kujul, mis tekivad valgurikaste vakuoolide veetustumisel. Kui aleurooniteradel pole märkimisväärset sisestruktuuri, nimetatakse neid lihtsateks. Liitterades paikneb amorfse valgu sees üks või mitu valgukristalli või globoidi. Globoid on ümar, varufosforit sisaldav keha. Valgukristallid paisuvad vees ja värvuvad rea värvainetega. Varuvalgud on lihtsa ehitusega ning need ei kuulu raku elusosa koostisse. Aleurooniterad esinevad sageli liblikõieliste (Fabaceae) idulehtedes ning kõrreliste (Poaceae) endospermi välimises, nn. aleuroonikihis.
Lipiidseteks varuaineteks on rasvad ja õlid. Rasvad on tavalistel temperatuuridel tahked, õlid aga vedelad. Tahkeid rasvu esineb taimedes harva, õlitilgad on seevastu väga levinud. Õlipalmi (Elaeis) seemnete endospermis on leitud ka niitjaid rasvakristalle.
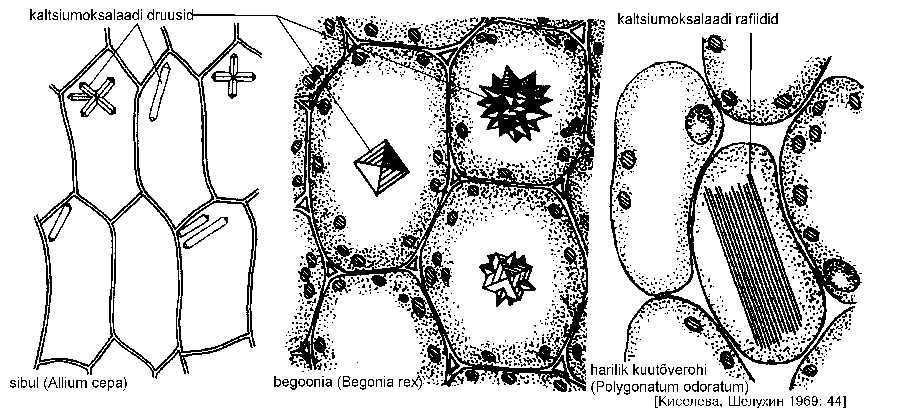
Ainevahetuse lõpp-produktidena eritatakse vakuoolidesse mitmesuguseid orgaanilisi happeid, millest üks tavalisemaid on oblikhape. See on rakkude ainevahetuse toksiline produkt, mis neutraliseeritakse Ca2+ ioonide abil -- moodustuvad kaltsiumoksalaadi (CaC2O4) kristallid. Kaltsiumoksalaat ladestub peamiselt vanades rakkudes druuside (liitoktaeedrid) või rafiididena (nõeljate kristallidena) (joonis). Druusid on iseloomulikud kaheidulehelistele, rafiidid aga üheidulehelistele taimedele, kuid esineb ka erandeid.
 Raku ainevahetuses
vajalikud ensüümid,
substraadid, kofaktorid jne.
peavad olema
kättesaadavad tsütoplasma
erinevates osades.
Väikestes rakkudes liiguvad
need ained ühest kohast
teise difusiooni teel.
Taimerakud on aga sageli
küllaltki suured ning sel juhul
osutub difusioon
väheefektiivseks. Siin aitab tsütoplasma voogav liikumine, mis on iseloomulik ainult elusatele
rakkudele. Kui tsütoplasma ringleb piki raku kesta, nimetatakse seda rotatsiooniks, kui aga
liikumine on nähtav plasmaväätides, siis tsirkulatsiooniks. Kuna tsütoplasma on värvitu,
näeme tsütoplasma liikumist organellide ja teiste rakusisaldiste liikumisena.
Raku ainevahetuses
vajalikud ensüümid,
substraadid, kofaktorid jne.
peavad olema
kättesaadavad tsütoplasma
erinevates osades.
Väikestes rakkudes liiguvad
need ained ühest kohast
teise difusiooni teel.
Taimerakud on aga sageli
küllaltki suured ning sel juhul
osutub difusioon
väheefektiivseks. Siin aitab tsütoplasma voogav liikumine, mis on iseloomulik ainult elusatele
rakkudele. Kui tsütoplasma ringleb piki raku kesta, nimetatakse seda rotatsiooniks, kui aga
liikumine on nähtav plasmaväätides, siis tsirkulatsiooniks. Kuna tsütoplasma on värvitu,
näeme tsütoplasma liikumist organellide ja teiste rakusisaldiste liikumisena.
Tsütoplasma liikumist käivitavaid mehhanisme on viimasel ajal molekulaarsel tasandil põhjalikult uuritud. Liikumise aluseks on aktiinifilamentide ja müosiinimolekulide omavaheline vastasmõju. Näiteks vetika Nitella rakkudes on tsütoplasma välimises kihis, kortikaalses tsütoplasmas, paiknevad aktiinifilamendid orienteeritud kõik sarnaselt. Müosiinimolekulidel aga on omadus ATP juuresolekul nihkuda aktiinifilamentide positiivse otsa suunas (ATP hüdrolüüsiga kaasneb müosiinimolekulide peas konformatsiooniline muutus, mis tekitab nihkumise aktiinifilamendi + otsa suunas). Müosiini libisemine mööda aktiinifilamente paneb ringlema kogu tsütoplasma. Aktiini-müosiini vastasmõju on ka mitmete teiste rakus toimuvate liikumiste (näit. lihaskontraktsioon, loomse raku tsütoplasma sissenöördumine tsütokineesi käigus jt.) käivitajaks.
Erinevalt loomsetest rakkudest ümbritseb taimerakke rakukest. Esmane rakukest koosneb valdavalt pektiinidest, hemitselluloosidest ning tselluloosist, on elastne ega takista rakkude kasvamist. Jäigemaks muudab selle mitmesuguste teiste ainete ladestumine. Paiguti on primaarne rakukest õhem; nendes kohtades (esmastel pooriväljadel) läbivad rakukesta plasmodesmide kanalid. Sekundaarse kesta kujunemisel tselluloos pooriväljadele ei ladestu, tekivad poorid. Rakukest pakseneb seestpoolt, s.t. primaarne kest jääb väljapoole, sekundaarne sissepoole.
Vanemate rakkude kestad enamasti puituvad (lignifitseeruvad) -- toimub ligniini (puitaine) ladestumine. See muudab rakukestad deformatsioonidele vastupidavaks. Rakukesta võib ladestuda ka mitmesuguseid mineraalaineid (CaCO3, SiO2). Mitmete puuliikide puit on nendest nii läbi imbunud, et löögi mõjul see ei lõhene, vaid puruneb kildudeks.
Rakukesta mehaaniline tugevus võimaldab taimerakul viibida hüpotoonilises keskkonnas (soolade kontsentratsioon väljaspool rakku on madalam kui raku sees). Taimerakku ümbritsev vedelik on alati hüpotoonilisem kui raku sisekeskkond. Seetõttu liigub vesi osmoosi läbi rakku ning tekib rõhk, mida nimetatakse turgoriks. Turgor annab suure osa taime rohtsete kudede mehaanilisest tugevusest. Hüpertoonilises lahuses (väljaspool rakku on soolade kontsentratsioon kõrgem) hakkab vesi osmoosi tõttu rakust välja liikuma kõrgema kontsentratsiooniga lahuse suunas, kusjuures rakukest etendab poolläbilaskva membraani osa. Toimub plasmolüüs -- tsütoplasma tõmbub veekaotuse tagajärjel kokku. Eristatakse osalist ja täielikku, nõgus ja kumerplasmolüüsi. Kui asetada osaliselt plasmolüüsunud rakk tagasi hüpotoonilisse keskkonda, toimub deplasmolüüs. See näitab, et rakk pole veel surnud, tegemist oli üksnes osalise plasmolüüsiga. Täielikult plasmolüüsunud rakk on surnud ja deplasmolüüsi enam ei toimu. Nõgus- ja kumerplasmolüüs erinevad plasmolüüsunud osa kuju poolest.
Rakke liidab rakuvaheaine (vahelamell), vanemate rakkude nurkade vahel võib olla ka rakuvaheruume ehk intertsellulaare.
Raku ehitusest paremaks arusaamiseks pidage silmas, et see pole mitte jäik, staatiline moodustis, vaid raku ehitus ja koostis muutuvad pidevalt ja küllaltki kiiresti: organellid jagunevad või kujunevad ümber, keemiline koostis muutub jne.
Koeks nimetatakse ühesuguse päritolu, ehituse ja talitlusega rakkude rühma. Ehituslikult lihtsaim on ainult üht tüüpi rakkudest koosnev lihtkude (näiteks kollenhüüm, sklerenhüüm). Liitkude on keerukam, kuna koosneb mitut tüüpi rakkudest (näiteks epiderm, floeem). Koes esinevaid üksikuid teistsuguse ehituse ja ülesannetega rakke nimetatakse idioblastideks. Neid võib leiduda ka lihtkoes.
Enamasti ei asu rakud tihedalt üksteise kõrval, vaid nende vahele jäävad suuremad või väiksemad rakuvaheruumid ehk intertsellulaarid. Kuigi rakuvaheruumid on eriti iseloomulikud vanadele või vähemalt väljaarenenud kudedele, on neid leitud ka algkudedest. Eriti suuri rakuvaheruume esineb lehtedes ning vee- ja sootaimede vartes, kus nende funktsiooniks on elutegevuseks ja vees püstipüsimiseks vajalike gaaside säilitamine. Intertsellulaarid on koe struktuurseks osaks, nende hulk ja suurus on koele iseloomulik.
Rakuvaheruumid tekivad enamasti skisogeenselt: rakkusid liitev, peamiselt pektiinist koosnev vahelamell laguneb ja rakukestade vahele tekib tühimik. Skisogeense rakuvaheruumi teke algab mitme raku kesta kokkupuutekohast ning laieneb seejärel kesta ülejäänud osadele. Sarnaselt kulgeb ka matseratsioon -- vahelamelli lagundamine keemiliste (nt. kroomhappega) või füüsikaliste (leotamine, keetmine) meetoditega, mille tulemusel on võimalik uurida vigastamata üksikrakke. Kudede looduslik matsereerumine toimub näiteks lihakate viljade valmimisel.
Lüsigeensele intertsellulaari tekkele eelneb rakkude suremine, seejärel lahustub rakukest ja osa protoplasmast. Vähem levinud on rakuvaheruumide reksigeenne kujunemine, sel juhul rakud algul purunevad ja siis surevad. Intertsellulaarid võivad moodustuda ka kombineeritult, näiteks kõigepealt skisogeenselt, pärast aga lüsigeenselt.
Kudede kirjeldamisel kasutatakse sageli mõisteid antiklinaalne ja periklinaalne jagunemine (joonis). Esimesel
juhul tekib vahesein taimeorgani välispinnaga risti
(perpendikulaarselt), teisel paralleelselt. Telgorganeid (vars,
varb, leheroots jt.) kirjeldades kasutatakse sageli antiklinaalse asemel radiaalse (vaheseinad tekivad telgorgani
kujutletava pikiteljega samas suunas ja organi välispinnaga
risti) ning transversaalse (vaheseinad kujunevad risti nii
pikiteljega kui ka organi välispinnaga) jagunemise terminit.
Periklinaalse jagunemise asemel räägitakse sel juhul
tangentsiaalsest. 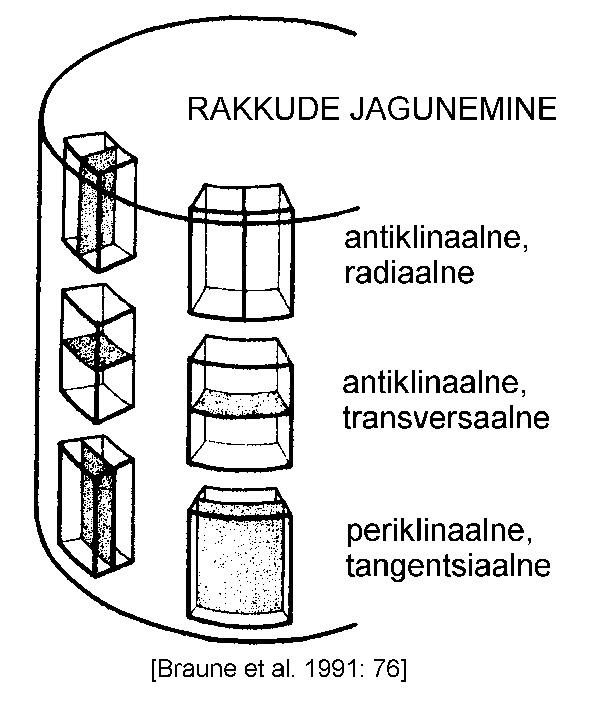
Ehkki kudede klassifitseerimisel pole seni ühtsele seisukohale jõutud (koetüübid on seotud üleminekutega, enamasti täidavad koed samaaegselt mitut ülesannet, sageli muutuvad nende funktsioonid taime ontogeneesi käigus), on kudede liigitamine vajalik võrdlevatel uuringutel, taimede ehituse ja ontogeneesi kirjeldamisel ning kudede mitmekesisuses orienteerumisel.
Uute organite teket diferentseerumata algkoest ehk meristeemist nimetatakse organogeneesiks. Sellele eelneb histogenees -- initsiaalrakkudest püsikudede moodustumine. Mõlemat protsessi koos nimetatakse morfogeneesiks.
Algkude koosneb pidevalt jagunevatest, piiramatu jagunemisvõimega initsiaalrakkudest ning edasises arengus püsikudedeks diferentseeruvatest initsiaalrakkude derivaatidest. Enamasti kaotavad initsiaalrakkude derivaadid (tugikoe, juhtkoe jms. rakud) paljunemisvõime küllaltki kiiresti, ainult põhikoe rakkudel säilib see omadus kauem.
Algkoe rakud on väikesed ja õhukese tselluloosse kestaga, sest kiire pooldumise tõttu ei jõua need kuigi palju kasvada. Rakutuumad on kogu raku mõõtmetega võrreldes suhteliselt suured, vakuoolid aga väikesed või puuduvad hoopis. Kujult on algkoe rakud hulktahksed (kasvukuhikus) või prosenhüümsed (näiteks kambiumis). Rakuvaheruumid on silmatorkamatud, rakud asuvad tihedalt üksteise kõrval.
Algkudesid, mis on taimes olemas tema arengu algusest peale, nimetatakse primaarseteks
meristeemideks ehk esimeristeemideks. Esimeristeemideks on ka lõigustuv sügoot
(viljastatud munarakk) ja idujuure ning idupunga tippudes paiknevad tipmised meristeemid.
Neist saavad alguse rest- ehk püsimeristeemid, nagu kambium, sporogeenne algkude või
meristemoidid (näiteks õhulõhede emarakud, vt. allpool). Mitmeaastastel taimedel võib aga
püsikoe rakkudest tekkida uus algkude, mida nimetatakse sekundaarseks ehk teismeristeemiks. Selline algkude on näiteks vahetult epidermi all (subepidermaalselt) paiknev,
esikoore põhikoest, aga ka kollenhüümist või epidermist tekkinud fellogeen ehk korgikambium, mille tegevusel tekib mitmeaastaste
kaheiduleheliste taimede vartele periderm (korgikiht) ja
korp. 
Algkudede liigitamisel lähtutakse sageli nende ruumilisest asukohast taimes (joonis). Nii eristatakse tipmisi ehk apikaalseid (varte ja juurte tippudes, pungades, neid nimetatakse ka kasvukuhikuteks), külgmisi ehk lateraalseid (kambium) ning vahe- ehk interkalaarseid (varre sõlmekohtades asuvaid) meristeeme.
Taime vigastatud koha ümber tekib haavakude ehk kallus, milles leidub algkoelisi rakke -- kalluse- ehk haavameristeemi. Selle talitledes tekivad teisesed haavakoed -- haavapuit, haavakork jne. või isegi uued organid. Ka kallusemeristeem on sekundaarne algkude.
Apikaalsed ehk tipmised algkoed asuvad varre ja juure tipus. Need algkoed paiknevad koonilisest moodustisena -- kasvukuhikuna, mida ümbritsevad soomusjad lehe primordiumid (lehealgmed).
Lihtsaima kasvukuhiku tipus on üks suur piiramatu pooldumisvõimega, kolmetahulisele püramiidile sarnanev kiird- ehk initsiaalrakk, mis jaguneb kordamööda kõikide külgede suunas ja moodustab niimoodi kiirdraku derivaate. Kiirdrakk on tugevasti vakuoliseerunud, kuid derivaatide protoplasmas on vakuoolid väikesed. Kirjeldatud kasvukuhiku tüüp esineb soontaimede alamatel esindajatel -- hõimkonnas sõnajalgtaimed (Pteridophyta).
Paljas- ja katteseemnetaimede kasvukuhiku tipus asub grupp piiramatu pooldumisvõimega initsiaalrakke. Initsiaalrakke ja nende esimesi derivaate pole kuigi hõlpus eristada, sest olulised morfoloogilised erinevused nende vahel puuduvad (joonis).
Initsiaalrakkude derivaatide edasist diferentseerumist seletatakse tavaliselt tuunika-korpus teooria järgi, mille esitas A. Schmidt 1924. aastal. Selle kohaselt koosneb kasvukuhiku initsiaaltsoon 1) tuunikast, mis moodustub ühest või mõnest antiklinaalselt jagunevast rakkude kihist ning 2) korpusest, mille rakud jagunevad mitmes erinevas suunas. Seega on korpus keskne jagunevate rakkude rühm, mida katab tuunika. Katteseemnetaimede enamikul esineb kahekihiline tuunika, kuid on leitud ka 4-5 kihilist. Piir tuunika ja korpuse vahel pole alati selgesti eristatav ning siis sõltub juba uurijast, kas ta vaidlustatavad kihid arvab tuunika või korpuse hulka. Paljasseemnetaimedel puuduvad stabiilselt antiklinaalselt jagunevad rakukihid ning tuunikat pole võimalik eristada.
Kirjeldatust erineb klassikaline, I. Hansteini esitatud histogeenide teooria, mille kohaselt
epiderm, esikoor ja kesksilinder kujunevad kasvukuhiku kindlatest initsiaalrakkudest
(dermatogeen, peribleem, pleroom). 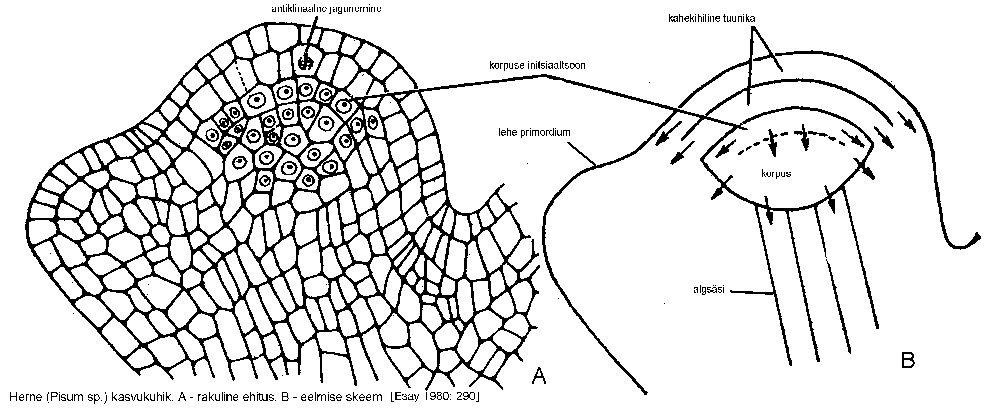
Kaheiduleheliste taimede teiskasv on põhjustatud lateraalse meristeemi -- kambiumi talitlusest. Kambium koosneb kahesugustest rakkudest: prismaatilistest prosenhüümsetest initsiaalrakkudest ning peaaegu isodiameetrilistest säsikiire initsiaalrakkudest. Säsikiire initsiaalrakud loovad säsielemente, prismaatilised aga floeemi ja ksüleemi. Teistest meristeemidest eristab kambiumi rohkete vakuoolide esinemine.
Vööndilisest kambiumist on põhjust kõnelda siis, kui prismaatilised rakud paiknevad tangentsiaallõigul selgesti eristuvate ridadena. Kambiumi initsiaalrakkude paigutus määrab puidu ehituse ning seega kujuneb vööndilise kambiumi esinemisel vastavalt ka vööndiline puit.
Floeemi- ja ksüleemirakkude moodustamisel jagunevad kambiumirakud periklinaalselt. Aktiivse kasvu perioodil toimub see nii kiiresti, et uue jagunemise korral jäävad eelmis(t)e jagunemis(t)e rakud meristemaatilisteks, need ei jõua diferentseeruda. Moodustub nn. kambiaalne tsoon, mida sageli käsitletakse kui mitmekihilist kambiumi. Kuigi pooldumisvõime on vaid initsiaalrakkudel, pole neid morfoloogiliselt võimalik eristada teistest kambiaalse tsooni rakkudest. Vahetevahel jagunevad kambiumirakud ka antiklinaalselt, mis tagab kambiumi püsimise pideva rõngana. Kambiumirakud toodavad ksüleemi umbes 4-10 korda rohkem kui floeemi, seetõttu on need kihid erineva tüsedusega.
Meristemoidideks nimetatakse üksikuid rakke või rakkude gruppe, millel on säilinud võime edasi diferentseeruda kindlateks struktuurideks (näiteks epidermis asuvad õhulõhe emarakud) ja mis paiknevad meristemaatilise aktiivsuse ammu kaotanud, väljakujunenud kudedes. Meristemoididele on sageli iseloomulik jagunemine kaheks ebavõrdse suurusega rakuks, millest üks on suur, kuid talitluslikult väheaktiivne ning teine on väike, füsioloogiliselt aktiivne ning hästiarenenud tsütoplasmaga. Selline nähtus ilmneb näiteks tolmutera vegetatiivse ja generatiivse raku, floeemi sõeltoru ja saateraku, õhulõhe sulg- ja kaasraku või juurekarvade moodustumisel.
Põhikoed moodustavad valdava osa taimede kehast. Kuna põhikoe rakud on kujult enamasti parenhüümsed, nimetatakse põhikudet tihti ka põhiparenhüümiks või lihtsalt parenhüümiks. Arusaamatuste vältimiseks tuleks siiski eelistada sisuliselt täpsemat mõistet "põhikude", tähistades sõnaga "parenhüüm" vaid rakkude tüüpi, sest ka teised koed võivad koosneda parenhüümsetest rakkudest.
Isegi siis, kui põhikoe rakud on peaaegu isodiameetrilised, pole need kujult kerajad, vaid hulktahksed. Homogeenses põhikoes peaksid rakud olema 14-tahksed kehad, kuid see ideaalkuju on tegelikkuses harvaesinev. Tahud puutuvad kokku naaberrakkude tahkudega, rakud on erineva suurusega, mistõttu enamasti on tahkusid 10-16. Põhikoerakud võivad olla ka tähtjad (vee- ja sootaimede aerenhüüm).
Põhikudede ühiseks omaduseks on võime muuta oma aktiivsust. Seda võimaldab kõikide organellide esinemine protoplastis. Põhikoe rakud osalevad regeneratsioonis ja lisajuurte ja -võsude moodustamisel. Vastavas toitekeskkonnas võib mõnest üksikust põhikoelisest rakust areneda terve taim.Funktsioonist lähtudes eristatakse järgmisi põhikoe liike.
Klorenhüüm (assimilatsioonipõhikude) asub lehes või varre esikoores. Üksikuid kloroplaste võib esineda ka säsis, kui sinna ulatuvad päikesekiired. Ometigi ei anna kloroplastide esinemine alust nimetata seda klorenhüümiks, sest fotosüntees pole sellise koe põhiülesandeks.
Klorenhüümi rakud on vakuoolide- ja kloroplastiderohked. Piklikud ja üksteise kõrval asuvad klorenhüümirakud moodustavad sammas- ehk palissaadkoe, kui aga rakkude vahel on suured rakuvaheruumid, nimetatakse seda kobe- ehk tohlkoeks.
Säilituspõhikoe rakkudesse kogunevad varuained: tärklis, lahustunud suhkrud, varuvalk (aleurooniterad) või -rasv (tilkadena). Viljades on sel juhul ka rohkesti kromoplaste. Varuainete säilitamise ülesandega rakkude kestad võivad viljades olla tugevasti paksenenud, nagu näiteks asparil (Asparagus), kohvipuul (Coffea arabica), diospüüril (Diospyros) või datlipalmil (Phoenix dactylifera). Sel juhul kasutab arenev idu rakukestades leiduvat hemitselluloosi.
Veesäilituskude esineb sukulentidel ehk turdtaimedel, näiteks sugukondades piimalillelised (Euphorbiaceae), kaktuselised (Cactaceae); perekondades agaav (Agave), kukehari (Sedum), soolarohi (Salicornia). Rakud on õhukesekestalised, paisumisvõimelised, veerohkete vakuoolidega. Kui vakuoolides on vett vähe, tõmbuvad rakkude kestad kortsu, rakkude veega täitudes sirguvad kestad uuesti.
Aerenhüüm (õhukude) sisaldab suuri rakuvaheruume, sest rakud on kujult tähtjad. Idioblastidena võib esineda tugikoerakke. Esineb soo- ja veetaimedel, siin on aerenhüümil oluline osa gaasivahetuses. Lisaks aitab aerenhüüm veetaimedel püsida vees vertikaalasendis.
Põhikudesid saab liigitada ka nende paiknemise järgi taimes:
esikoore põhikude (esikoore parenhüüm) asub epidermi ja kesksilindri vahel, selle rakud on vähespetsialiseerunud;
säsi on varre keskosas paiknev vähe spetsialiseerunud põhikude;
ksüleemi- ja floeemipõhikude asub enamasti säsikiirtena juhtkimpude floeemi ja ksüleemi juhteelementide vahel. Ksüleemipõhikoe rakud võivad puituda ning seetõttu väliselt sarnaneda sklerenhüümile, mis raskendab nende eristamist tugikoe rakkudest.
Eristatakse välimisi kattekudesid, mille funktsiooniks on katta taime välispinda (kaitse kahjulike välismõjude eest, transpiratsiooni ja gaasivahetuse regulatsioon) ning sisemisi kattekudesid, nagu epiteel vaigukanalite seintel või juure kesksilindrit ümbritsev endoderm. Sisemised kattekoed reguleerivad ainete liikumist taime sees.
Kasvukuhikust tekkinud kattekoed on esmased ehk primaarsed, sekundaarsetest meristeemidest tekkinud kattekoed aga teisesed ehk sekundaarsed. Primaarsetest kattekudedest on kõige sagedam epiderm, mis esikasvu (primaarse kasvu) staadiumis katab kogu taime keha. Juure epidermi nimetatakse epibleemiks ehk risodermiks. Samuti on esmasteks kattekudedeks lehtedes vahetult epidermi all asuv hüpoderm ja juure esikoore välimine kiht -- eksoderm ning troopiliste epifüütide juurenahk (velamen radicum). Eksoderm on ühe või mõne rakukihi paksune osadel juurtel esinev kattekude.
Teiskasvu (sekundaarkasvu) käigus asendub nii varte epiderm kui ka juurte epibleem peridermi ehk korgikihiga. Selle nimetusega tähistatakse kõiki fellogeenist ehk korgikambiumist alguse saanud kudesid. Fellogeen tekib kas epidermist, esikoore põhikoest, floeemist või peritsüklist. Väljapoole toodab fellogeen felleemi ehk korkkudet, sissepoole õhukese kihi fellodermi ehk korgi põhikudet.
Tugikoed (mehaanilised koed), mida sageli peetakse põhikudede alaliigiks, hoiavad või aitavad hoida taime keha püsti. Tugikoed puuduvad või on vähe arenenud veetaimedel ja sageli ka üheaastastel maismaataimedel. Veetaimi hoiab vertikaalasendis gaaside üleslükkejõud ning vee suur tihedus, noortel ja üheaastastel taimedel piisab püstipüsimiseks turgorist ehk rakkude siserõhust.
Tugikoed jaotatakse ehituslike, talitluslike ja geneesi iseärasuste alusel kahte põhitüüpi -- kollenhüümiks ja sklerenhüümiks.
Kollenhüüm on lihtsaim tugikude, mis koosneb ebaühtlaselt paksenenud kestadega elusatest, enamasti prosenhüümsetest rakkudest. Kollenhüüm on primaarne tugikude. Selle rakukestad ei puitu, vaid koosnevad tselluloosist, pektiinidest ja hemitselluloosist ning ei takista seega taime kasvu. Kollenhüümirakud on elastsed, heledad ja veerikkad, kuna pektiinid on hüdrofiilsed.
Küllaltki põhjendatud on kollenhüümi arvamine hoopis põhikudede hulka. Ka kollenhüümi rakud võivad olla meristemaatiliselt aktiivsed ja moodustada näiteks fellogeeni või haavakoe. Oluliseks erinevuseks on kollenhüümi rakukestade suurem paksus ja rakkude suurem pikkus (kuni 2 mm) põhikoe rakkudega võrreldes, kuid kollenhüümi ja põhikoe kokkupuuteala piir on sageli ebaselge.
Kollenhüüm esineb telgorganites vahetult epidermi all pideva rõngana või ribadena (joonis). Lehelaba suuremaid roode (juhtkimpe) ümbritseb paksenenud kestadega põhikude, mida samuti nimetatakse vahel kollenhüümiks. See kude erineb oma tekkelt ja struktuurilt tüüpilisest kollenhüümist, mis esineb üksnes varte perifeersetes osades. Seetõttu on õigem nimetada lehtedes esinevat "kollenhüümi" kollenhümiseerunud põhikoeks. Leherootsus paikneb kollenhüüm samuti nagu varreski, juures esineb väga harva.
Kollenhüümi tüüpe eristatakse rakukestade paksenemise iseloomu järgi (joonis: A-C):
1. Nurkkollenhüüm -- rakukestad paksenevad vaid nurkadest. Levinuim kollenhüümi tüüp.
2. Plaatkollenhüüm -- paksenevad vaid telgorgani välispinnaga paralleelsed (tangentsiaalsed) rakukestad, radiaalsed jäävad õhukesteks. Vähelevinud, esineb näiteks tähtputke (Astrantia) leherootsus ja musta leedri (Sambucus nigra) aastavõrses.
3. Kobekollenhüüm --
paksenevad
rakuvaheruumidega kokku
puutuvad rakukestade
piirkonnad. Näib, nagu
oleks suurte rakkude vahel
väiksemad paksukestalised
rakud, mis aga tegelikult on
rakuvaheruumid. 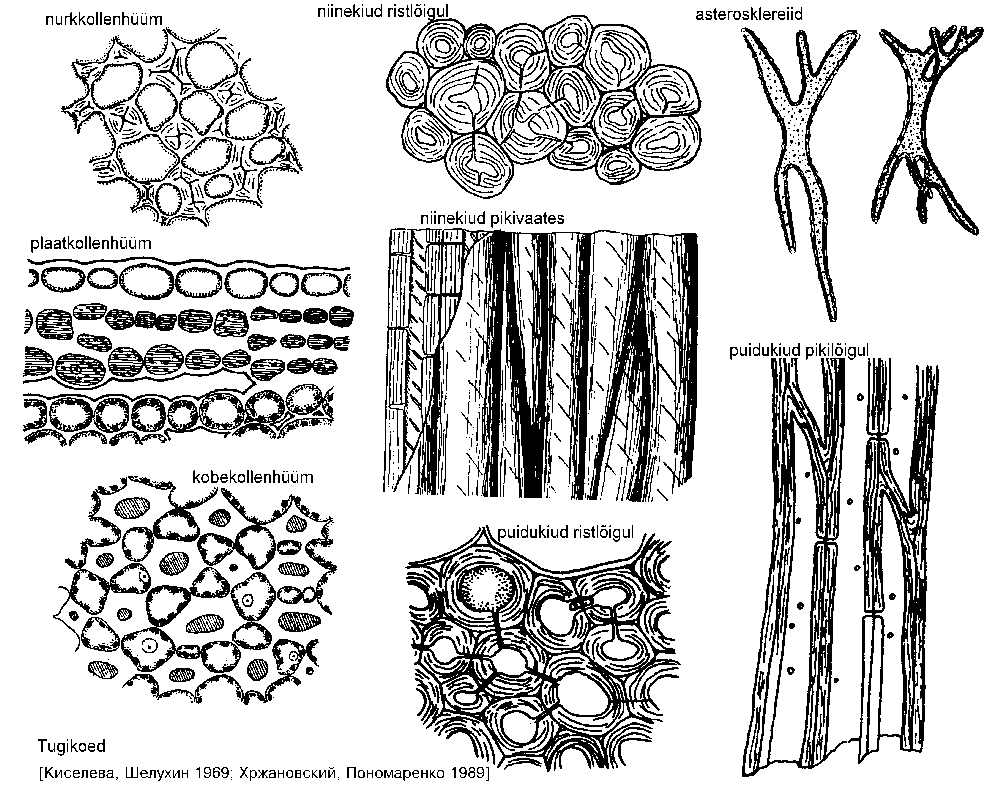
Kollenhüümi rakukestadele ladestub pidevalt uusi pektiini ja hemitselluloosi kihte. Seega muutuvad taime vananemisel erinevused kollenhüümi tüüpide vahel üha väiksemaks, seda tuleb arvestada koe identifitseerimisel. Näiteks nurkkollenhüümi iseloomulikud nurgapaksendused on sageli omavahel seotud paksenenud rakukestadega. Taime edasises arengus võivad kollenhüümi rakkude kestad puituda ning seega tekkida sklerenhüüm. Vanades taimedes leidub kollenhüümi lehtedes ja rohtsetes vartes, kus sklerenhüüm on vähearenenud.
Sklerenhüüm on sekundaarne tugikude, mis enamasti
koosneb teritunud otstega prosenhüümsetest rakkudest.
Asub varre siseosas (joonis). Täiskasvanult on sklerenhüümirakud surnud ja õhuga täitunud, rakukestad on
enamasti puitunud, vaid üksikutel liikidel jäävad need
tselluloosseteks. Sklerenhüümi hulka kuuluvad sklereiidid,
niine- ja puidukiud. 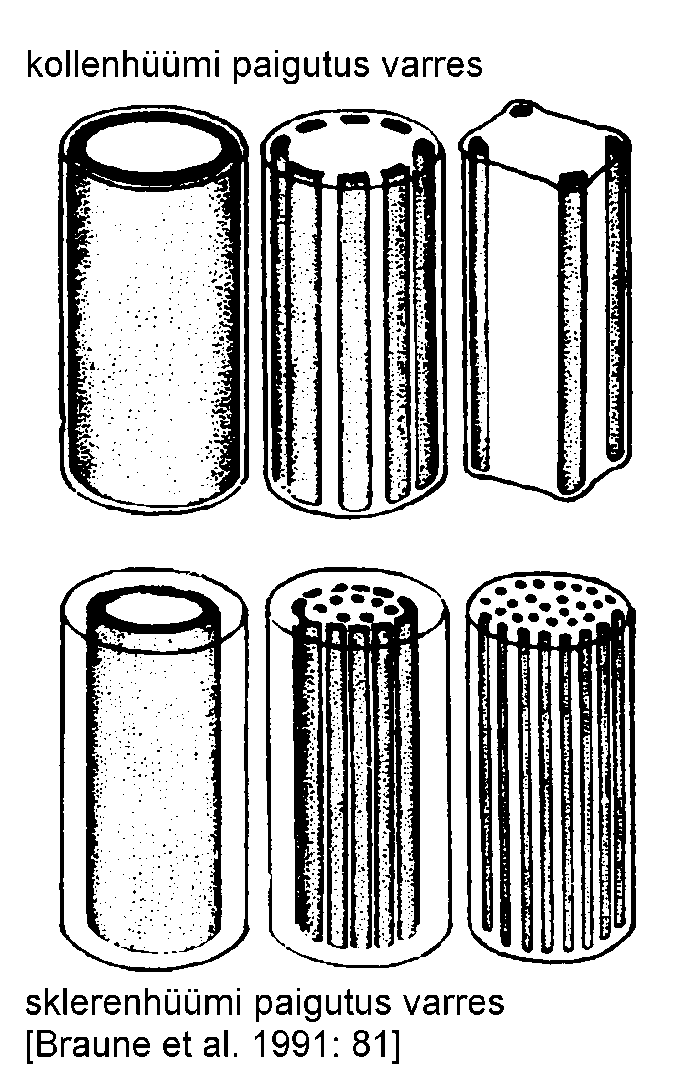
1. Sklereiidide ehk kivisrakkude kestad on tugevasti puitunud, pooridega läbistatud ja silmatorkavalt kihilised. Sklereiide võib leida idioblastidena põhikoes, kuid need võivad moodustada ka väga tugeva koe näiteks luuviljaliste seemnes või sarapuupähklis. Kivisrakkude kuju varieerub tugevasti ning sellel põhinevad klassifikatsioonid on sagedaste üleminekute tõttu vähekasutatavad. Brahhiosklereiidid on isodiameetrilise ümara kujuga kivisrakud, asterosklereiidid (joonis: F) seevastu meenutavad kujult tähte. Prosenhüümseid kivisrakke nimetatakse makrosklereiidideks. Kui need on kujult otsekui otstest laienenud toruluud ning moodustavad pideva kihi näiteks seemne- või viljakestas, nimetatakse neid osteosklereiidideks. Harunenud taimekarvadele sarnanevad trihhosklereiidid ning niinekiududele niitjad sklereiidid. Ka viimased võivad olla harunenud.
2. Niinekiud (joonis: D, E) on otstest teritunud, väga paksude kestadega rakud. Esinevad kas üksikult või kimpudena juhtkimpude floeemiosas ning kserofüütide lehtedes. Tekivad kas algkoest (primaarsed niinekiud) või kambiumist (sekundaarsed niinekiud). Niinekiudude pikkus ulatub mõnest millimeetrist 20-40 mm-ni või enam, kuid nende läbimõõt on vaid 0,1-0,2 mm.
Leidub kaheidulehelisi taimi, kelle kesksilindri perifeerses osas esineb niinekiudusid, mis ei kuulu floeemi koostisse, vaid asuvad sellest väljaspool. Kui neid on vaja eristada floeemsetest niinekiududest, võib neid nimetada peritsüklilisteks niinekiududeks, kuna need kujunevad peritsükli funktsioneerides. Üksikutel liikidel, näiteks perekonnas lina (Linum), jäävad niinekiudude kestad puitumata. Esineb ka pisut puitunud (kanep -- Cannabis) või täiesti puitunud (pärn -- Tilia) niinekiude.
 3. Puidukiud (libriformikiud, joonis: G, H) asuvad harilikult
juhtkimpude ksüleemis. Sarnanevad niinekiududele, kuid on
rohkem puitunud ja lühemad, kuni 1,5 mm pikkused. Esinevad
peaaegu kõikides mitmeaastastes taimedes. Okaspuudel
harilikult puuduvad, lehtpuupuidus esinevad.
3. Puidukiud (libriformikiud, joonis: G, H) asuvad harilikult
juhtkimpude ksüleemis. Sarnanevad niinekiududele, kuid on
rohkem puitunud ja lühemad, kuni 1,5 mm pikkused. Esinevad
peaaegu kõikides mitmeaastastes taimedes. Okaspuudel
harilikult puuduvad, lehtpuupuidus esinevad.
Ainete transport taime maapealsete ja maa-aluste organite vahel toimub mööda juhtkudesid. Juurte kaudu siseneb vesi koos selles lahustunud mineraalainetega taime ning liigub edasi temamaapealsetesse osadesse. Seda nimetatakse tõusvaks vooluks ning vastavat juhtkimbu osa, mida mööda vesi liigub, ksüleemiks ehk puiduosaks. Laskuv vool, mis kannab orgaanilised ained lehtedest vartesse ja juurtesse, samuti õitesse ning viljadesse, kulgeb floeemiks ehk niineosaks nimetatava juhtkimbu piirkonna kaudu. Nii ksüleem kui ka floeem läbib katkematult kogu taime (joonis).
Floeem koosneb sõeltorudest, saaterakkudest, floeemipõhikoest (floeemiparenhüümist) ja niinekiududest, mõnedel liikidel lisandub nimetatutele idioblastidena veel sklereiide jt. rakke (joonis).
Sõeltorud koosnevad piklikest, enamasti tömpide otstega prosenhüümsetest rakkudest. Nende tipmised ja külgmised rakukestad on varustatud rohkete pooridega, mis on koondunud pooriväljadeks. Katteseemnetaimede sõeltorude otstes on pooride läbimõõt tunduvalt suurem kui külgseinte pooriväljadel ning taolist raku otsmist poorivälja nimetatakse sõelplaadiks.
Seni kuni sõeltoru funktsioneerib, on tema rakud elusad. Tsütoplasma on õhukese kihina liibunud vastu rakukesta, siseruumi täidab suur vakuool. Rakutuum hävib varakult. Vähehaaval hakkab sõeltorude pooride ümber kogunema eriline aine -- kalloos, mille tagajärjel poorid lõpuks ummistuvad ning rakud surevad. Üheidulehelistel funktsioneerivad sõeltorud kogu taime eluea vältel. Enamikul kaheidulehelistest elavad sõeltorud ühe, vaid üksikutel liikidel mitu aastat.
Saaterakud esinevad peaaegu kõigil katteseemnetaimedel. Need on pikad kitsad elusad rakud, mis paiknevad sõeltorude kõrval. Enamasti arenevad need sõeltoruga samast algkoe rakust. Naaberrakkude protoplasmad on omavahel ühendatud plasmodesmide abil.
Sõeltorusid, saaterakke ja floeemipõhikude kui floeemi
elusat, juhtkoena talitlevat osa nimetatakse
leptoomiks. 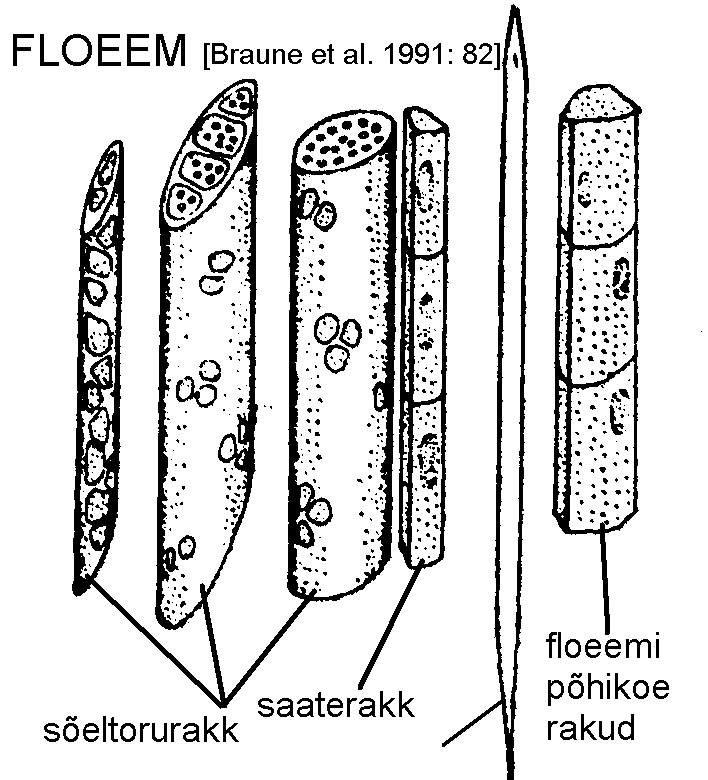
Ksüleemi funktsiooniks on vee ja mineraalainete transport, varuainete säilitamine ja taime toestamine. Ksüleemis võivad esineda trahheiidid, trahheed, ksüleemi põhikude ja puidukiud (joonis).
Trahheiidid esinevad enam-vähem kõikidel soontaimedel. need on prosenhüümsed, tippudest ahenenud, täiskasvanult surnud, puitunud kestadega rakud. Trahheiidide rakukesta siseküljel võib olla mitmesuguseid spiraalseid või rõngasjaid paksendeid. Paksendite iseloomu alusel eristatakse spiraal-, rõngas-, astrik- ja poortrahheiide. Trahheiidi sees liiguvad lahused takistamatult nagu torus ikka, kuid ühest rakust teise pääseb vedelik läbi trahheiidikesta pooride. Lihtsamal juhul kujutavad poorid endast sulgkilega varustatud kanalit, ehituselt keerukamad on koobaspoorid.
 Trahheed (sooned) on trahheiididest täiuslikumad. need esinevad enamikul katteseemnetaimedel ning vähestel sõnajalg- ja paljas-seemnetaimedel. Trahhee rakkude otsmised
rakukestad on kas kadunud, nii et moo-dustub
pidev toru, või on rakukestadesse tekkinud
piklikud avad. Ka trahheed on täiskasvanult
surnud ja puitunud, nende rakukestad võivad
seestpoolt olla spiraalsete (spiraaltrahhee),
rõngasjate (rõngastrahhee) või võrkjate
(võrktrahhee) paksendustega. Astriktrahhee
kestas asetsevad korrapäraste ridadena piklikud
poorid, rakkude tipud on kolmnurksed ja piklike
pooridega läbistatud. Täiuslikuim trahhee tüüp on
poortrahhee, mille külgmistes rakukestades on
koobaspoorid, otsmised kestad on alati hävinud.
Poortrahhee läbimõõt võib ulatuda millimeetrini.
Trahheed (sooned) on trahheiididest täiuslikumad. need esinevad enamikul katteseemnetaimedel ning vähestel sõnajalg- ja paljas-seemnetaimedel. Trahhee rakkude otsmised
rakukestad on kas kadunud, nii et moo-dustub
pidev toru, või on rakukestadesse tekkinud
piklikud avad. Ka trahheed on täiskasvanult
surnud ja puitunud, nende rakukestad võivad
seestpoolt olla spiraalsete (spiraaltrahhee),
rõngasjate (rõngastrahhee) või võrkjate
(võrktrahhee) paksendustega. Astriktrahhee
kestas asetsevad korrapäraste ridadena piklikud
poorid, rakkude tipud on kolmnurksed ja piklike
pooridega läbistatud. Täiuslikuim trahhee tüüp on
poortrahhee, mille külgmistes rakukestades on
koobaspoorid, otsmised kestad on alati hävinud.
Poortrahhee läbimõõt võib ulatuda millimeetrini.
Puidu põhikude koosneb nõrgalt puitunud kestadega elusatest põhikoerakkudest, mis paiknevad kitsaste radiaalsete ribadena -- säsikiirtena. Need rakud on varuainete säilituskohaks ja samal ajal kindlustavad ainete liikumise radiaalsuunas.
Puidukiud esinevad vaid katteseemnetaimede puidus, okaspuudes neid pole.
Trahheiide ja trahheesid koos puidu põhikoega nimetatakse hadroomiks.
Primaarne ksüleem ja primaarne floeem diferentseeruvad prokambiumist juba varakult. Primaarse ksüleemi kõige väiksema valendikuga ja algelisematest rõngas- ja spiraaltrahheiididest koosnevat osa nimetatakse protoksüleemiks. Hiljem tekib metaksüleem, mis koosneb suurematest spiraal-, astrik- ja poortrahheiididest. Meta- ja protoksüleemi vaheline piir on enamasti raskesti jälgitav, kuna erinevused nende vahel on väikesed. Teiskasvu esinedes arenevad kimbukambiumi tegevuse tagajärjel sekundaarne floeem ja sekundaarne ksüleem.
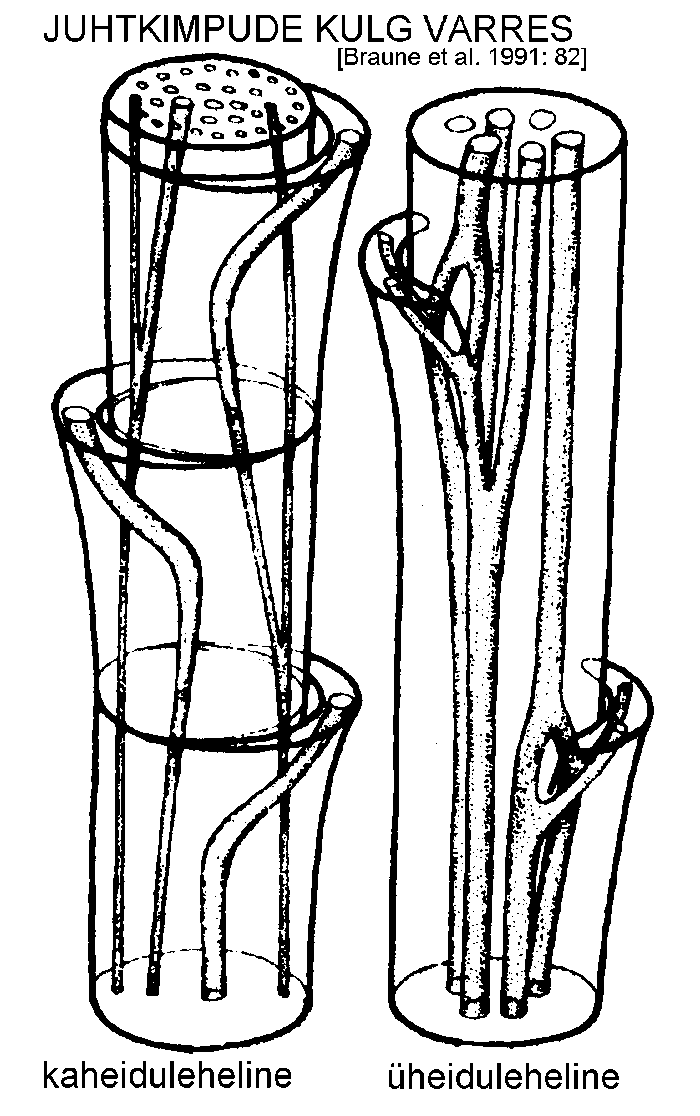
Floeem ja ksüleem ühes tugikoe, kambiumi ja põhikoe elementidega moodustavad juhtkimbu. Üheidulehelistel ja osadel kaheidulehelistel taimedel puudub juhtkimbus kambium ning juhtkimp säilitab kogu taime eluea jooksul algse ehituse. Sellist ehitustüüpi nimetatakse kinniseks juhtkimbuks.
Kaheidulehelistel ja enamikul paljasseemnetaimedel esineb kambium, seega ka teiskasv, ning juhtkimp on avatud. Kui juhtkimp koosneb kas ainult floeemist või ksüleemist, räägitakse mittetäielikust juhtkimbust. Enamikul veetaimedel esinevad redutseerunud juhtkimbud, mille ksüleem on vähearenenud. Redutseerunud juhtkimpe leidub ka maismaataimede lehtedes.
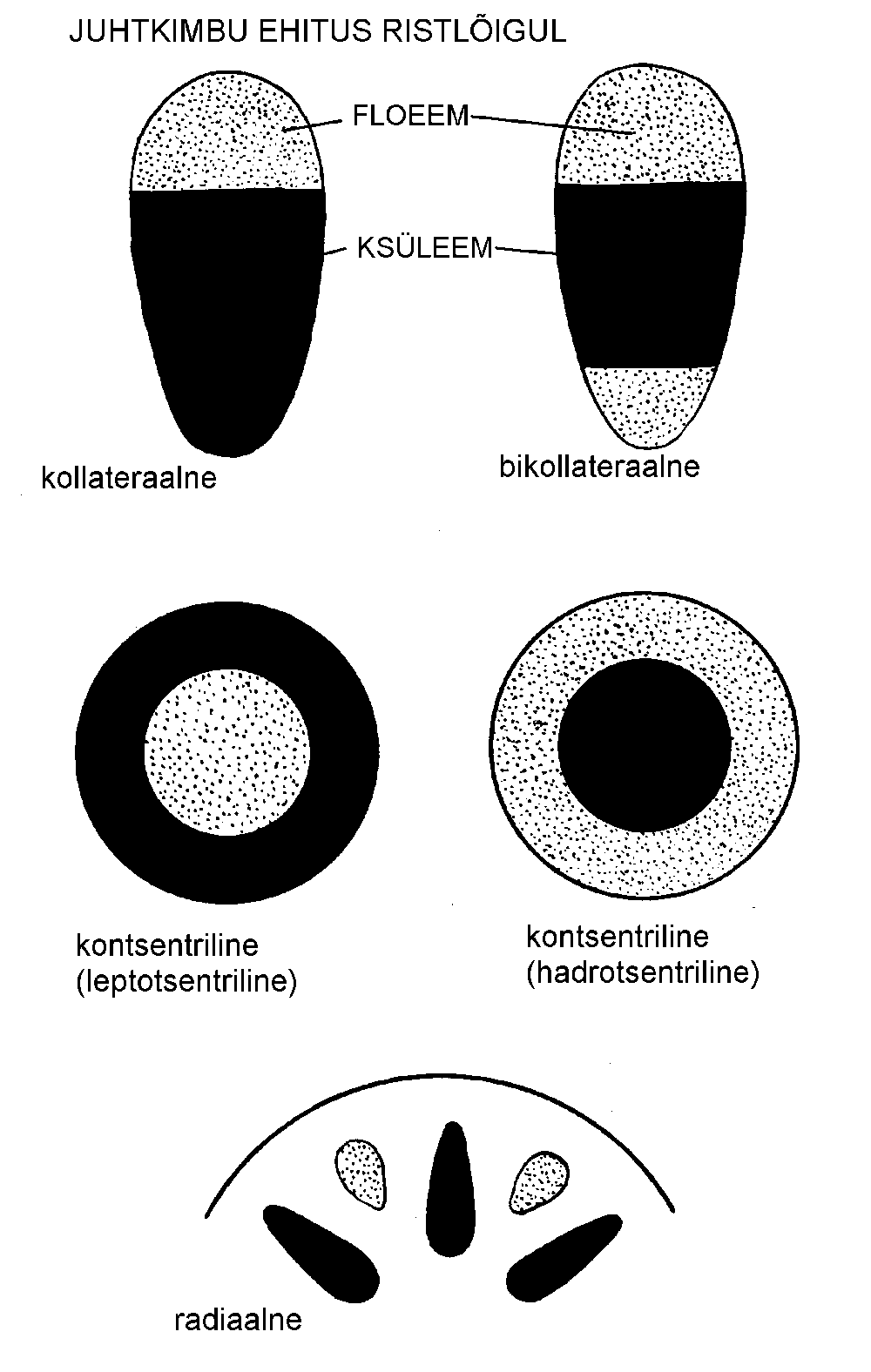
Floeemi ja ksüleemi paigutuse järgi eristatakse järgmisi juhtkimpude tüüpe (joonis):
kontsentriline juhtkimp. Kui floeem on ksüleemist täielikult ümbritsetud, räägitakse leptotsentrilisest, kui aga vastupidi, siis hadrotsentrilisest juhtkimbust. Kontsentrilised juhtkimbud esinevad sagedamini risoomides;
kollateraalne juhtkimp. Ksüleem ja floeem paiknevad kõrvuti, floeem enamasti juhtkimbu välispinna pool, ksüleem aga seespool. Sagedaseim juhtkimbu tüüp;
bikollateraalne juhtkimp. Floeem asetseb kahel pool ksüleemi. Esineb korvõieliste (Asteraceae), maavitsaliste (Solanaceae), kõrvitsaliste (Cucurbitaceae) jt. sugukonnas;
radiaalne juhtkimp. Ksüleem ja floeem paiknevad vaheldumisi. Radiaalne juhtkimp on iseloomulik juurtele.
Enamasti ümbritseb juhtkimpu sklerenhüümist või kollenhümiseerunud põhikoest koosnev, ühe- või mitmerakukihiline kimbutupp.
 Erituskudede ülesandeks on mitmesuguste ainete eritamine
kas vedelal või tahkel kujul. Kui eritatav aine on taime
elutegevuseks vajalik, nimetatakse seda sekreediks
(nõreks), kui aga mittevajalik, siis ekskreediks (eritiseks).
Niisugune jaotus on üsna tinglik, sest teadmised ühe või
teise aine (näiteks kautđuk, eeterlikud õlid, alkaloidid jt.)
osast taime ainevahetuses ja elutegevuses on sageli lünklikud. Allpool on eritamise ehk sekretsiooni mõistet käsitletud
laialt, lugedes siia nii ainete eritamise rakkudest väliskeskkonda kui ka taimes olevaisse mahuteisse.
Erituskudede ülesandeks on mitmesuguste ainete eritamine
kas vedelal või tahkel kujul. Kui eritatav aine on taime
elutegevuseks vajalik, nimetatakse seda sekreediks
(nõreks), kui aga mittevajalik, siis ekskreediks (eritiseks).
Niisugune jaotus on üsna tinglik, sest teadmised ühe või
teise aine (näiteks kautđuk, eeterlikud õlid, alkaloidid jt.)
osast taime ainevahetuses ja elutegevuses on sageli lünklikud. Allpool on eritamise ehk sekretsiooni mõistet käsitletud
laialt, lugedes siia nii ainete eritamise rakkudest väliskeskkonda kui ka taimes olevaisse mahuteisse.
A. Eritatavad ained jäävad taime. Selliselt talitlevateks erituskudedeks on lülilised ja lülitud piimasooned, osa idioblaste, mitmesugused mahutid ja käigud.
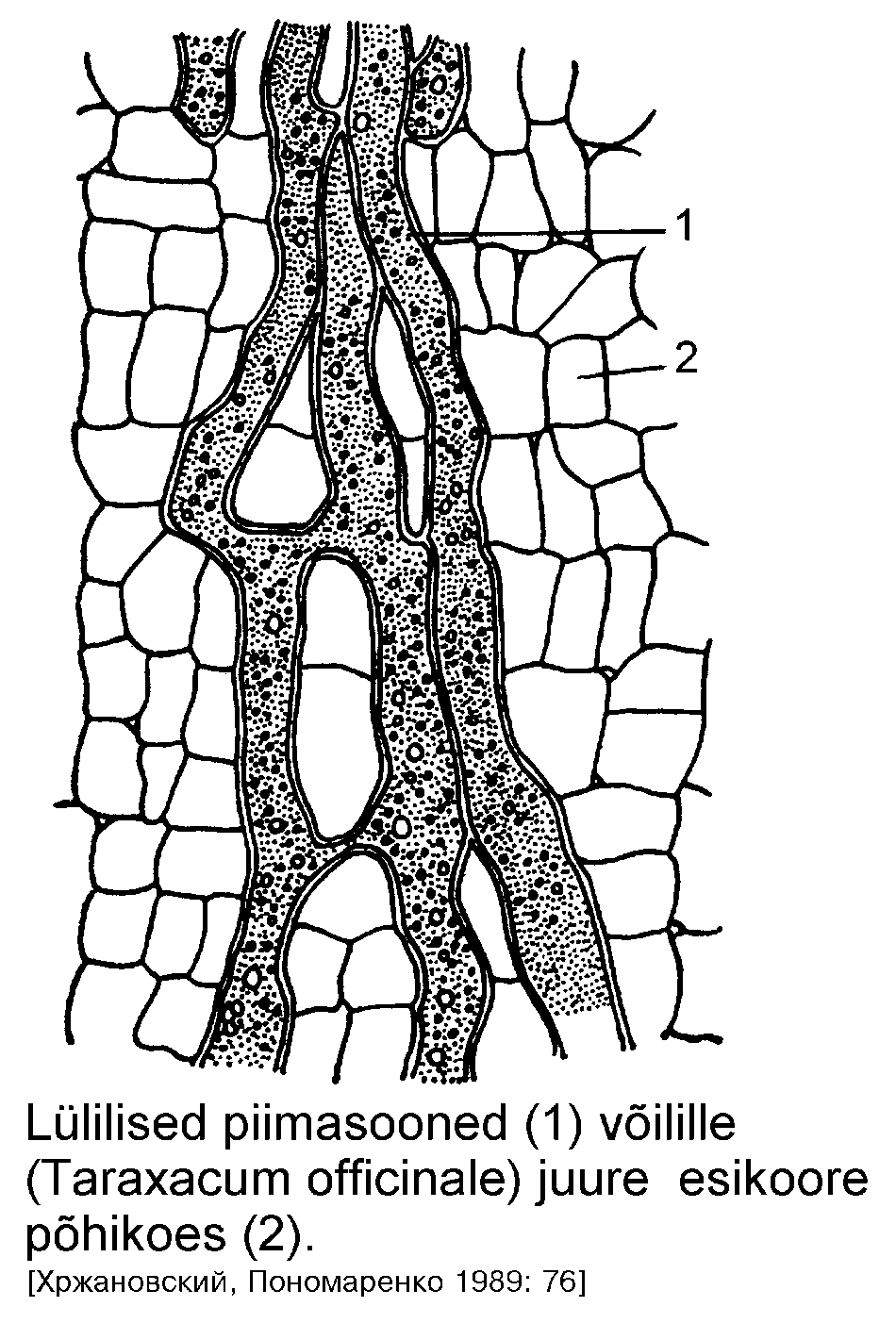
Lülilised piimasooned on pikad torujad kanalid, mis tekivad sarnaselt trahheedele või sõeltorudele: rakud liituvad otstest ja otsmised rakukestad hävivad. Kui piimasoonte vahel esinevad anastomoosid ehk ühendused, moodustub kanalite võrgustik. See on iseloomulik näiteks korvõieliste alamsugukonnale keelõielised (Liguliflorae, joonis) ja magunatele (Papaver). Anastomoosideta piimasooni esineb näiteks perekondades kassitapp (Convolvulus), lauk (Allium) ja vereurmarohi (Chelidonium).
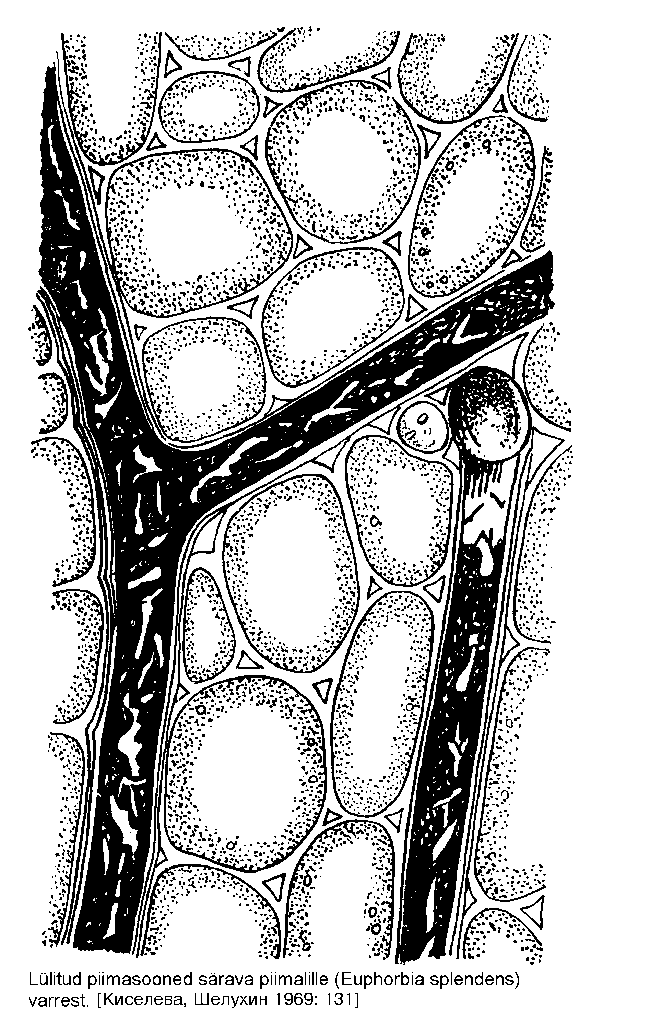
Lülitute piimasoonte rakud ei liitu, vaid iga piimasoon koosneb pikast, torujast rakust. Selliseid rakke ei tarvitse ühes taimes olla palju (5-6), kuid raku pikkus võib ulatuda viie meetrini. Harunevad lülitud piimasooned on perekondades igihali (Vinca), nõges (Urtica), kanep (Cannabis); harunematud esinevad perekondades piimalill (Euphorbia, joonis), oleander (Nerium), viigipuu (Ficus) jt. Nii lüliliste kui lülitute piimasoonte funktsiooni suhtes puudub botaanikutel ühine seisukoht.
Idioblastid on sageli eritusfunktsiooniga. Nende produktideks on mitmesugused limad, eeterlikud õlid, kaltsiumoksalaadi kristallid (rafiidide või druusidena), alkaloidid, parkained jms.
 Mahutid ja käigud erinevad sekretoorsetest idioblastidest selle poolest, et kujutavad endast lüsigeenselt või
skisogeenselt tekkinud õõnsust.
Mahutid ja käigud erinevad sekretoorsetest idioblastidest selle poolest, et kujutavad endast lüsigeenselt või
skisogeenselt tekkinud õõnsust.
Lüsigeensed sekretoorsed mahutid tekivad sekreeti sisaldavate rakkude hävimisel. Need esinevad näiteks ruudiliste (Rutaceae) sugukonna perekonnas tsitrus (Citrus) või kassinaeriliste (Malvaceae) perekonnas puuvill (Gossypium).
Skisogeensed sekretoorsed õlimahutid esinevad eukalüptidel (Eucalyptus, sugukond mürdilised, Myrtaceae). Need tekivad ühe epidermiraku järglastest, mis diferentseeruvad epiteeliks ja piirderakkudeks. Edasises arengus kujuneb epiteelirakkude vahele skisogeenne mahuti. Samalaadse tekkega on ka okaspuude vaigukäigud, pärnaliste (Tiliaceae) sugukonna limakäigud, korvõieliste (Asteraceae) tõrvakäigud jne.
B. Ained eritatakse väliskeskkonda. Siia kuuluvad epidermaalsed näärmed ja hüdatoodid.
Epidermaalsed näärmed, mis koosnevad üksikutest rakkudest ja meenutavad väliskujult trihhoome, on näärmekarvad. Kui rakke on rohkem ja nääre ei meenuta karvu, siis nimetatakse seda näärmeepiteeliks.
Tähtsaimaks näärmeepiteeli tüübiks on nektaariumid. Nende funktsiooniks on suhkruid sisaldavate vedelike eritamine. Esinevad enamasti õites (floraalsed nektaariumid), harvem väljaspool õisi (ekstrafloraalsed nektaariumid). Õites paiknevad need tupp- või kroonlehtedel, tolmukatel, sigimikul või õiepõhjal, ekstrafloraalseid nektaariume võib leida vartelt, lehtedelt, abilehtedelt ja õieraagudelt. Nektaariumid on sageli juhtkimpude läheduses või on neil omaette, sageli vaid floeemist koosnev juhtkimp.
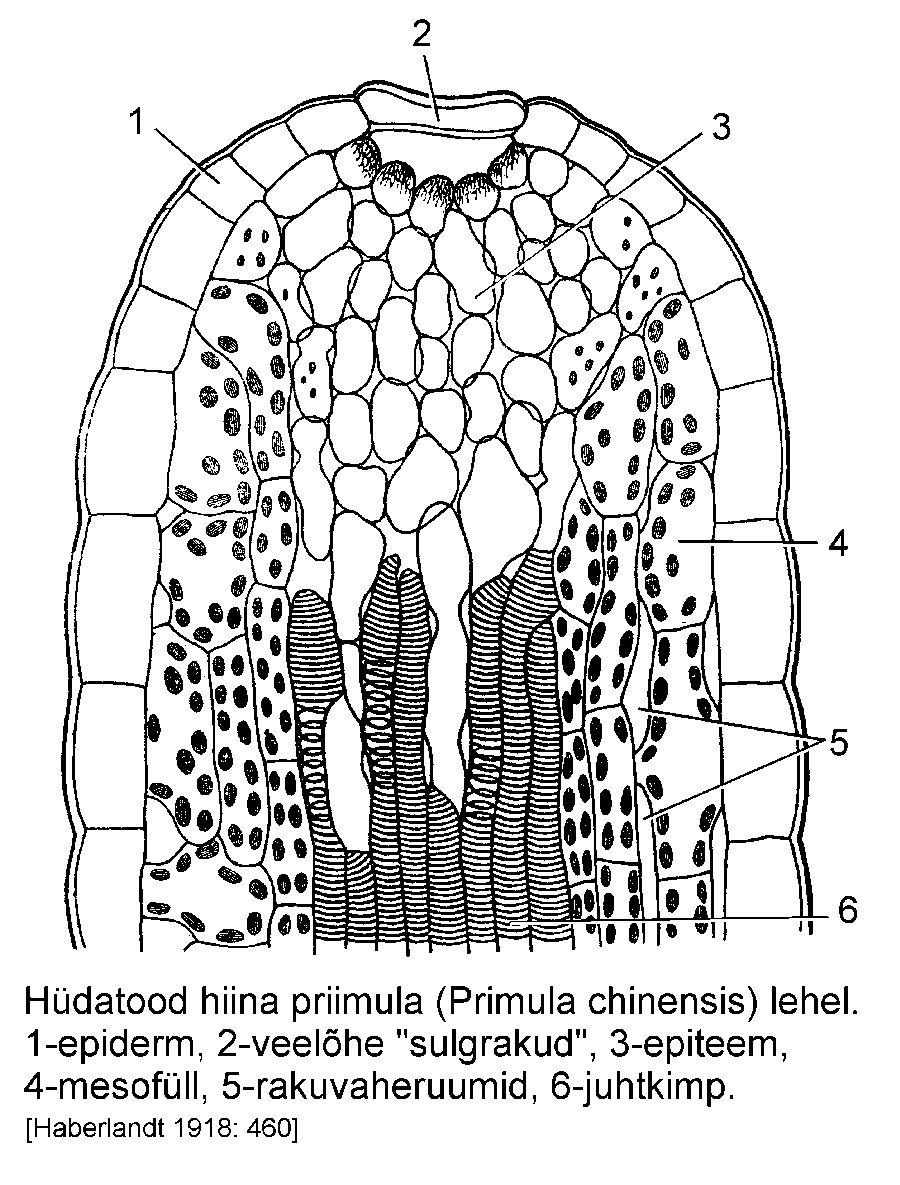 Hüdatoodid (veelõhed) eritavad lehe välispinnale
vett. Ehituslikult meenutavad need õhulõhesid
(joonis). Lehe välispinnal või servas on näha kaks
piklikku rakku, mille vahele jääb veelõhe. Taim ei
saa hüdatoodi avada ega sulgeda. Selle all on
hulgaliselt õhukese kestaga kloroplastideta rakke
(modifitseerunud mesofüll), mis moodustavad erilise
veeerituskoe -- epiteemi. Hüdatoode on mitmesuguse suuruse ja keerukusega. Ühed hüdatoodid
eritavad vett passiivselt juurerõhu mõjul, teised on
näärmelise epiteemiga, mis on võimeline vett
eritama juurerõhust sõltumatult. Vee eritamist
nimetatakse ka gutatsiooniks.
Hüdatoodid (veelõhed) eritavad lehe välispinnale
vett. Ehituslikult meenutavad need õhulõhesid
(joonis). Lehe välispinnal või servas on näha kaks
piklikku rakku, mille vahele jääb veelõhe. Taim ei
saa hüdatoodi avada ega sulgeda. Selle all on
hulgaliselt õhukese kestaga kloroplastideta rakke
(modifitseerunud mesofüll), mis moodustavad erilise
veeerituskoe -- epiteemi. Hüdatoode on mitmesuguse suuruse ja keerukusega. Ühed hüdatoodid
eritavad vett passiivselt juurerõhu mõjul, teised on
näärmelise epiteemiga, mis on võimeline vett
eritama juurerõhust sõltumatult. Vee eritamist
nimetatakse ka gutatsiooniks.
 2.5.3.
2.5.3.